Structural Biochemistry/Virus Entry by Endocytosis
Overview of virus entry
editEven though viruses are not complex and quite simple in terms of their structure and their components, the way they interact with host cells are quite complex. In order to enter the cell, animal viruses utilize a wide variety of cellular processes that deal with numerous cellular proteins. Even though some viruses are able to go into the cytosol through the plasma membrane, the majority of the viruses rely on endocytic uptake, vesicular transport via the cytoplasm, and the transportation to the endosomes and the other intracellular organelles. This process of taking in the viruses are associated with clathrin-mediated endocytosis, macropinocytosis, caveolar/lipid raft-mediated endocytosis, or other mechanisms. There are many ways and endocytic mechanisms that animal viruses utilize to allow a virus to enter a cell.

Introduction of virus entry
editInitially viruses attach to the cell surface of proteins, carbohydrates, and lipids. The interactions virus receptors usually are specific and have at least 3 valences. In addition, these interactions result in the activation of cellular signaling pathways. Cells react by incorporating the viruses utilizing a few endocytic mechanisms. After the viruses go in the lumen of endosomes or the endoplasmic reticulum, they obtain signals which are in the form of being exposed to low pH, proteolytic cleavage, and the initiation of viral proteins. These cause modifications in the viral proteins, and then they are able to penetrate the vacuolar membrane. After they penetrate the vacuolar membrane, they pass the viral genome, the caspid, or the viral particle that is kept together into the cytosol. Afterwards, the majority of RNA viruses replicate at a variety of positions within the cytosol. In contrast, most DNA viruses continue through their passage towards to the nucleus. A process that consists of steps that dissemble and uncoat operate parallel with the movement of the virus and viral capsids deeper into the cell. This whole process results in a regulated dispersion of the genome and accessory proteins in a replication-competent form.
It is important to study and understand the virus-host cell interactions for a variety of reasons. Number one, the threat and potential damage that pathogens and viruses can cause are growing exponentially due to many factors like the world population growing larger, the immense increase in international trade and travel, and global warming. Therefore, it is extremely important and helpful to obtain as much information that can assist in the fight against viruses that are currently present or viruses that can potentially come up in the future. The study of the interactions between viruses and the cell is a growing field with many undiscovered details that can assist in developing strategies to deal with viruses.
Another important reason to study the interaction between viruses and cells is that viruses can be utilized as instruments in molecular medicine. Viruses have developed to the point where they can enter cells and bring in foreign genes and macromolecules. This is why they are very useful in gene therapy and transporting macromolecules and drugs into cells. In addition, they viruses of have the capacity to perhaps be able to pinpoint cancer cells and exterminate them. Viruses are an ever increasing important instrument that scientists can utilize and manipulate to develop new strategies and concepts of in the field of molecular, structural and cell biology.
Advantages of entry through endocytosis
editMorphological, genetic, and biochemical studies of bacteriophages have shown that the mechanisms of infection are extremely complicated. They also showed that the formation of coliphages T4 and T2 are made like hypodermic syringes with a tail that is capable of contracting. This creates a machinery that has the ability to strike through the two membranes of a gram-negative bacteria and penetrate the cytosol to send in the DNA. It was discovered that attachment to host cell receptors have the function of being a signal that activates the process of injection. Because the host cells of animal cells do not have the outer membrane and cell wall, they do not require complex and elaborate tools and mechanism to enter the cell. In contrast, bacteria has these cell wall and membranes that inhibit the pathway to the plasma membrane. In addition, animal cells give endocytic mechanisms that send viruses that come in advantages that bacteriophages do not possess. Endocytic vesicles carry viruses that come in from the outer edges to the perninuclear area of the host cell, where the conditions for infection are encouraged and the distance is minimized towards the nucleus. In effect, this enables viruses to cross through obstacles freely that deal with cytoplasmic bunching up and the complex construction of microfilaments in the cortex. The ability to travel through the endocytic vesicles is especially crucial for viruses that infect neurons, where the separate axons from the cell body have very long distances. Also the maturation of endosomes that have slowly changing conditions, like lowering of pH or the switching of a redox environment, enables viruses to detect their position within a cell and the passage. This will also allow the endosomes to utilize this information to put a time of penetration and uncoating. When certain proteases like furin and cathepsins are present they give the required proteolytic activation of speciic viruses. In addition, when animals viruses are endocytosed, they are able to avoid leaving evidence of them being exposed on the plasma membrane.It is important to avoid leaving evidence of them being exposed on the plasma membrane because then it would result in a delay in being detected by immunosurveillance. Combined as one, endocytosis contains numerous advantages that cause viruses, like herpes simplex virus 1 and human immunodefiency virus 1 to utilize endocytic pathways for productive entrance rather than entering directly.
The study of viral endocytosis has been combined with reviews on endocytoic pathways. It is important understand both of these aspects in these topics of wide range.
Viruses as Endocytic Carriers
editAnimal viruses have sizes that range from approximately 30 nm for parvoviruses to 40 nm for poxviruses. The majority of viruses lie in the 60 to 150 nm range. Even though viruses are typically shaped like a sphere, there are other viruses like Filovirus, Paramyxovirus, and influenza virus that can be fibrous and very stretched apart. When they attach to cells, viruses do not become disfigured. On the other hand, the plasma membrane changes shape usually to adjust to the shape of the virus. There are a few instances were invagination is necessary for endocytosis. The outer layer of viruses is usually covered proteins that attach to receptors in the form of capsid proteins formed in an icosahedral grid or as spike glycoprotein that span the entire viral envelope. The separate interactions with the receptors are usually weak, but interaction with many different receptor cause the activity to increase really high which causes the binding to cells almost impossible to reverse. Multivalent binding causes receptors to cluster, which may cause an association with lipid domains and activation of signaling pathways.
At the time when virus particles arrive at the endosomes, the virus particles are usually similar in size to the intralumenal vesicles. When they are too large to fit into the narrow tubular extensions, they are usually localized to the bulbous, vacuolar areas of the endosomes and are organized to the degradative pathway.
Viruses usually in the past have been utilized as model cargo in endocytosis and membrane trafficking studies. By using an electron microscopy one can easily recognize these viruses and they can be identified with fluorescent groups or proteins, which enable single-particle location and monitoring in live cells. By giving a focused source of light, the center of mass of fluorescent viruses can be accurately outlined using a point-spread function. Because infection causes amplification, when viruses have successfully entered the cell, it can be easily measured, even with very small amounts of virus. Also the many studies of virology has created numerous tools, like virus mutants, fluorescent viruses, antibodies, expression systems, and modified host cells.
Lipid raft function to control the signaling, fluidity and receptor functions on the membrane. These are usually rich in cholesterol and sphingolipids. Viruses that use the Caveolar/Raft-dependent pathways form primary endocytic vesicles that are dependent on cholesterol, lipid rafts, and complex signaling pathways. These ways use tyrosine kinases, phosphatases, and glycophingolipids. The process is started by the ligands and the site of penetration for these viruses is the ER. One of the most studied virus that uses this pathway is the polyomavirus. These viruses make use of the multiple receptors available in order to bind better.
Attachment Factors and Receptors
editEven though some viruses utilize receptors that have defined endocytic receptor functions, like transferrin and low-density lipoprotein receptors, the majority of molecules that viruses attach to deals with different functions like cell to cell recognition, ion transpotation, and attachment to the extracellular matrix. Most of the time the carbohydrate moieties serve as a important function in the binding of viruses.
It is important and helpful to distinguish the difference between attachment factors that simply attach viruses and therefore assist them to focus the viruses on the top area of the cell, and virus receptors that function as a trigger to modify the viruses, promote cellular signaling, or activate penetration. One objective that a lot of receptors have in common is that they encourage endocytosis and they assist the virus into the cell. A lot of the time, when the viruses enter, they begin with binding to attachment factors. Afterwards they interact with other receptors. In reality, it is hard to distinguish the difference between attachment factors and receptors because both of them play a role in the effectiveness of the infection.
The attachment factor that is most seen is called glycosaminoglycan chains in proteogylcans. Attaching to these negatively charged polysaccharides is typically electrostatic and somewhat not specific. There has been identification that in some instances viruses evolve to utilise GAGs when they adjust to the development in tissue culture. Another common category of carbohydrates consist of sialic acids, to which many viruses attach to. Similar to the situation with influenza and polyomaviruses, the majority of the time, this attachment is very specific and deals with lectin domains or lectin sites.
Because binding is very specific, this is a significant cause in determining tropism and species specificity and therefore one can tell the nature of viral diseases. One other dimension of receptor specificity is that it can identify the choice of endocytic pathway and intracellular routing that the viruses there are coming in will travel through. In this specific instance, parvoviruses that bind to the transferrin receptor utilize clathrin-mediated uptake passage. They are also capable to recycle to cell surface with their receptor. In contrast, not as significantly present groups like rhinoviruses that bind to the LDL receptor break apart from the receptor in early endosomes and taken to the late endosomes.
There are a few receptors that have the job of promoting modifications the virus that allows attachment of the virus to a coreceptor, promotion of endocytic uptake, or transformation to configuration that is membrane fusion-active. The most suitable defined case is HIV-1. This is because in this case two receptors are necessary to promote conformational modifications to promote the fusion. Adenoviruses 2 and 5 contain two receptors that promote conformational modifications and encourage endocytosis. For the avian leukosis virus, the signals necessary to entrance are receptor binding in conjunction with low levels of pH.
How viruses hijack cell regulation SLiMs
editHow viruses hijack cell regulation SLiMs Virus' life cycle is shaped by the interactions they have with cellular proteins. They can take over and use proteins for their own interest for their mechanisms. However, viruses are compact spatial constrained with tiny genomes, yet they can command so many pathways and processes. One of the reason they are able to do so is by extensive mimicry of host protein short linear motifs (SliMs).
-
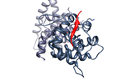 The Human papilloma virus E7 oncoprotein mimic of the LxCxE motif (red) bound to the host Retinoblastoma protein (dark grey)(PDB 1gux.
The Human papilloma virus E7 oncoprotein mimic of the LxCxE motif (red) bound to the host Retinoblastoma protein (dark grey)(PDB 1gux.

When viruses enter the cell, they have their own DNA and RNA polymerases, helicases and proteases, which enable replication, expression, and maturation. But, the virus still needs the cell to help with viral replication. Compact non globular protein interaction interfaces known as SliMs are present in eukaryotic proteomes. They have a wide variety of specific functions, such as the ability to encode a functional interaction interface in a short sequence, participation in a region of proteins, and capacity to function independently, and convergently evolve. Their functions include targeting proteins to specific destinations, acting as recognition sites for proteolytic cleavage, and provide specific modifications., signaling, recognition, rewiring pathways, molecular switching.
There are a diverse set of host pathways targeted and their functionality. Some of them include the host target 14-3-3 by viral protein Rep68 with Virus AAV with the Motif RsxSxP. Another example is host target ALIX by viral protein Gag for HIV with motif LYPxxxL. Thus, there are many different mechanisms by how viruses use the motifs.
One of the mechanism is cellular transport. Directing proteins to destinations require large complexes or cytoskeletal structure of micro tubules. Consequently, viral proteins use the vast cellular infrastructure by copying the motifs. For example, signal mimicry is the driving force of the essential regulatory protein Rev of HIV.
Another mechanism is signal transduction. SliMs have low binding affinities and fast off-rates, which promotes fast reaction in response to variation stimuli. Thus, viruses copies the signaling motifs and deregulate their pathways.
A 3rd mechanism is controlling cellular protein levels. Viral proteins play roles as scaffolds, by redirecting substrate specificity to unconventional host protein targets. For example, in ubiquity ligage recruitment, the virus hijacks the cell resulting in polyubiquitylation of the host, followed by proteasonal degradation. By doing so, the cellular protein is altered, which enables the viral proteins to survive and replicate their genome. Destruction motifs can also be used to inhibit the destruction of the proteins as well, which can prolong the lives of pro viral proteins.
A 4th mechanism is transcriptional regulation, which is a very tightly controlled cellular process. Even so, even its regulation can be degraded by motifs mimicry.
A 5th mechanism is to modify the host proteome by deregulation of endogenous modification processes. Viral proteins can act as scaffolds between host enzymes and viral targets. A single modification can ultimately result in modifying an entire genome.
A 6th mechanism is their own modification. Viral proteins can also direct their own modification by mimicking docking and modification sites of host enzymes.
Viruses must use motifs because the regular function of the cell is largely controlled by SliMs. They are evolution's results to rewire cellular regulatory pathways, which makes them equally vulnerable to the faster evolving viral proteins, allowing the viral proteins to deregulate the pathways.
SliMs length, which are 3-10 residues long, allows it to convergently evolve. Usually, only one or two mutations are necessary for the creation of a new motif. RNA viruses usually go through 10^-5 – 10^-3 mutations in each position in a generation, while DNA viruses usually go through 10^-8 – 10^-5. Convergent evolution of these viral motifs can be examined through viral strains and isolations. Phenotype variations of the viral proteins are due to the adaption of motif specificity.
The size of viral genomes are largely restricted, yet they still interact extensively with the host cell. Once the viral protein acquires the interaction with the protein interface, it alters a pre-folded region, but the host cannot operate under multiple binding sites that are close to each other, so they do not have the ability to interact with a number of these distinct partners because they are spatially restricted. Viral proteomes usually have folded structures. Generally, a globular interface is not needed for the viral to take over a process, but nevertheless, multiple contrasting mechanisms are available to manipulate a specific pathway or protein.
Eukaryote regulatory systems are robust, due to the cellular networks cooperation. Viruses can also use the mechanisms to also be robust. The small sizes of motifs also enable viral proteins to be robust against the mutations and adaptions. Mimicry of SliMs are also robust.
Reference
editMercer, Jason, Mario Schelhaas, and Ari Helenius. "Virus Entry by Endocytosis." Annual Review of Biochemistry 79.1 (2010): 803-33.