This page was imported and needs to be de-wikified. Books should use wikilinks rather sparsely, and only to reference technical or esoteric terms that are critical to understanding the content. Most if not all wikilinks should simply be removed. Please remove {{dewikify}} after the page is dewikified.
The history of biochemistry spans approximately 400 years. Although the term “biochemistry” seems to have been first used in 1882, it is generally accepted that the word "biochemistry" was first proposed in 1903 by Carl Neuberg, a German chemist.
Biochemistry is the study of chemical processes in living organisms. Biochemistry governs all living organisms and living processes. By controlling information flow through biochemical signalling and the flow of chemical energy through metabolism, biochemical processes give rise to the incredible complexity of life. Much of biochemistry deals with the structures and functions of cellular components such as proteins, carbohydrates, lipids, nucleic acids and other biomolecules although increasingly processes rather than individual molecules are the main focus. Over the last 40 years biochemistry has become so successful at explaining living processes that now almost all areas of the life sciences from botany to medicine are engaged in biochemical research. Today the main focus of pure biochemistry is in understanding how biological molecules give rise to the processes that occur within living cells which in turn relates greatly to the study and understanding of whole organisms.
Among the vast number of different biomolecules, many are complex and large molecules (called polymers), which are composed of similar repeating subunits (called monomers). Each class of polymeric biomolecule has a different set of subunit types. For example, a protein is a polymer whose subunits are selected from a set of 20 or more amino acids. Biochemistry studies the chemical properties of important biological molecules, like proteins, and in particular the chemistry of enzyme-catalyzed reactions.
The biochemistry of cell metabolism and the endocrine system has been extensively described. Other areas of biochemistry include the genetic code (DNA, RNA), protein synthesis, cell membrane transport, and signal transduction.[1]
As early as the late 18th century and early 19th century, the digestion of meat by stomach secretions[2] and the conversion of starch to sugars by plant extracts and saliva were known. However, the mechanism by which this occurred had not been identified.[3]
In the 19th century, when studying the fermentation of sugar to alcohol by yeast, Louis Pasteur came to the conclusion that this fermentation was catalyzed by a vital force contained within the yeast cells called "ferments", which were thought to function only within living organisms. He wrote that "alcoholic fermentation is an act correlated with the life and organization of the yeast cells, not with the death or putrefaction of the cells."[4]
In 1878 German physiologist Wilhelm Kühne (1837–1900) coined the term enzyme, which comes from Greek "in leaven", to describe this process. The word enzyme was used later to refer to nonliving substances such as pepsin, and the word ferment used to refer to chemical activity produced by living organisms.
In 1897 Eduard Buchner began to study the ability of yeast extracts to ferment sugar despite the absence of living yeast cells. In a series of experiments at the University of Berlin, he found that the sugar was fermented even when there were no living yeast cells in the mixture.[5] He named the enzyme that brought about the fermentation of sucrose "zymase".[6] In 1907 he received the Nobel Prize in Chemistry "for his biochemical research and his discovery of cell-free fermentation". Following Buchner's example; enzymes are usually named according to the reaction they carry out. Typically the suffix -ase is added to the name of the substrate (e.g., lactase is the enzyme that cleaves lactose) or the type of reaction (e.g., DNA polymerase forms DNA polymers).
Having shown that enzymes could function outside a living cell, the next step was to determine their biochemical nature. Many early workers noted that enzymatic activity was associated with proteins, but several scientists (such as Nobel laureate Richard Willstätter) argued that proteins were merely carriers for the true enzymes and that proteins per se were incapable of catalysis. However, in 1926, James B. Sumner showed that the enzyme urease was a pure protein and crystallized it; Sumner did likewise for the enzyme catalase in 1937. The conclusion that pure proteins can be enzymes was definitively proved by Northrop and Stanley, who worked on the digestive enzymes pepsin (1930), trypsin and chymotrypsin. These three scientists were awarded the 1946 Nobel Prize in Chemistry.[7]
This discovery that enzymes could be crystallized eventually allowed their structures to be solved by x-ray crystallography. This was first done for lysozyme, an enzyme found in tears, saliva and egg whites that digests the coating of some bacteria; the structure was solved by a group led by David Chilton Phillips and published in 1965.[8] This high-resolution structure of lysozyme marked the beginning of the field of structural biology and the effort to understand how enzymes work at an atomic level of detail.[9]
Santorio Santorio in his steelyard balance, from Ars de statica medecina, first published 1614
The term metabolism is derived from the Greek Metabolismos for "change", or "overthrow".[10] The history of the scientific study of metabolism spans 400 years. The first controlled experiments in human metabolism were published by Santorio Santorio in 1614 in his book Ars de statica medecina.[11] This book describes how he weighed himself before and after eating, sleeping, working, sex, fasting, drinking, and excreting. He found that most of the food he took in was lost through what he called "insensible perspiration".[12]
Since then, biochemistry has advanced, especially since the mid-20th century, with the development of new techniques such as chromatography, X-ray diffraction, NMR spectroscopy, radioisotopic labelling, electron microscopy and molecular dynamics simulations. These techniques allowed for the discovery and detailed analysis of many molecules and metabolic pathways of the cell, such as glycolysis and the Krebs cycle (citric acid cycle). One of the most prolific of these modern biochemists was Hans Krebs who made huge contributions to the study of metabolism.[13] He discovered the urea cycle and later, working with Hans Kornberg, the citric acid cycle and the glyoxylate cycle.[14][15][16]
In 1960, the biochemist Robert K. Crane revealed his discovery of the sodium-glucose cotransport as the mechanism for intestinal glucose absorption.[17] This was the very first proposal of a coupling between the fluxes of an ion and a substrate that has been seen as sparking a revolution in biology.Today, the findings of biochemistry are used in many areas, from genetics to molecular biology and from agriculture to medicine.[18]
Bioenergetics is the part of biochemistry concerned with the energy involved in making and breaking of chemical bonds in the molecules found in biological organisms.
Growth, development and metabolism are some of the central phenomena in the study of biological organisms. The role of energy is fundamental to such biological processes. The ability to harness energy from a variety of metabolic pathways is a property of all living organisms. Life is dependent on energy transformations; living organisms survive because of exchange of energy within and without.
In a living organism, chemical bonds are broken and made as part of the exchange and transformation of energy. Energy is available for work (such as mechanical work) or for other processes (such as chemical synthesis and anabolic processes in growth), when weak bonds are broken and stronger bonds are made. The production of stronger bonds allows release of usable energy.[19]
Living organisms obtain energy from organic and inorganic materials. For example, lithotrophs can oxidize minerals such as nitrates or forms of sulfur, such as elemental sulfur, sulfites, and hydrogen sulfide to produce ATP. In photosynthesis, autotrophs can produce ATP using light energy. Heterotrophs must consume organic compounds. These are mostly carbohydrates, fats, and proteins. The amount of energy actually obtained by the organism is lower than the amount present in the food; there are losses in digestion, metabolism, and thermogenesis.
The materials are generally combined with oxygen to release energy, although some can also be oxidized anaerobically by various organisms. The bonds holding the molecules of nutrients together and the bonds holding molecules of free oxygen together are all relatively weak compared with the chemical bonds holding carbon dioxide and water together. The utilization of these materials is a form of slow combustion. That is why the energy content of food can be estimated with a bomb calorimeter. The materials are oxidized slowly enough that the organisms do not actually produce fire. The oxidation releases energy because stronger bonds have been formed (corresponding to lower energy). This net energy may evolve as heat, or some of which may be used by the organism for other purposes, such as breaking other bonds to do chemistry.
Living organisms produce ATP from energy sources via oxidative phosphorylation. The terminal phosphate bonds of ATP are relatively weak compared with the stronger bonds formed when ATP is broken down to adenosine monophosphate and phosphate, dissolved in water. Here it is the energy of hydration that results in energy release. This hydrolysis of ATP is used as a battery to store energy in cells, for intermediate metabolism. Utilization of chemical energy from such molecular bond rearrangement powers biological processes in every biological organism.[20]
The concept of entropy is defined by the second law of thermodynamics, which states that the entropy of a closed system always increases or remains constant.
Entropy change
When an ideal gas undergoes a change, its entropy may also change. For cases where the specific heat doesn't change and either volume, pressure or temperature is also constant, the change in entropy can be easily calculated.[22]
When specific heat and volume are constant, the change in entropy is given by:
.
When specific heat and pressure are constant, the change in entropy is given by:
.
When specific heat and temperature are constant, the change in entropy is given by:
.
In these equations is the specific heat at constant volume, is the specific heat at constant pressure, is the ideal gas constant, and is the number of moles of gas.
For some other transformations, not all of these properties (specific heat, volume, pressure or temperature) are constant. In these cases, for only 1 mole of an ideal gas, the change in entropy can be given by[23] either:
Enthalpy is a measure of the total energy of a thermodynamic system. It includes the internal energy, which is the energy required to create a system, and the amount of energy required to make room for it by displacing its environment and establishing its volume and pressure.
Note that the U term is equivalent to the energy required to create the system, and that the pV term is equivalent to the energy which would be required to "make room" for the system if the pressure of the environment remained constant.
The pV term may be understood by the following example of an isobaric process. Consider gas changing its volume (by, for example, a chemical reaction) in a cylinder, pushing a piston, maintaining constant pressure p. The force is calculated from the area A of the piston and definition of pressure p = F/A: the force is F = pA. By definition, work W done is W = Fx, where x is the distance traversed. Combining gives W = pAx, and the product Ax is the volume traversed by the piston: Ax = V. Thus, the work done by the gas is W = pV, where p is a constant pressure and V the expansion of volume. Including this pV term means that during constant pressure expansion, any internal energy forfeited as work on the environment does not affect the value of enthalpy. The enthalpy change can be defined ΔH = ΔU + W = ΔU + Δ(pV), where ΔU is the thermal energy lost to expansion, and W the energy gained due to work done on the piston.[25]
In thermodynamics, the Gibbs free energy (IUPAC recommended name: Gibbs energy or Gibbs function; also known as free enthalpy[1] to distinguish it from Helmholtz free energy) is a thermodynamic potential that measures the "useful" or process-initiating work obtainable from an isothermal, isobaric thermodynamic system.
The Gibbs free energy, originally called available energy, was developed in the 1870s by the American mathematician Josiah Willard Gibbs. In 1873, Gibbs described this “available energy” as the greatest amount of mechanical work which can be obtained from a given quantity of a certain substance in a given initial state, without increasing its total volume or allowing heat to pass to or from external bodies, except such as at the close of the processes are left in their initial condition.[26]
To derive the Gibbs free energy equation for an isolated system, let Stot be the total entropy of the isolated system, that is, a system that cannot exchange heat or mass with its surroundings. According to the second law of thermodynamics:
and if ΔStot = 0 then the process is reversible. The heat transfer Q vanishes for an adiabatic system. Any adiabatic process that is also reversible is called an isentropic process.
Now consider systems, having internal entropy Sint. Such a system is thermally connected to its surroundings, which have entropy Sext. The entropy form of the second law applies only to the closed system formed by both the system and its surroundings. Therefore a process is possible if
.
If Q is heat transferred to the system from the surroundings, so −Q is heat lost by the surroundings
so that corresponds to entropy change of the surroundings.
We now have:
Multiply both sides by T:
Q is heat transferred to the system; if the process is now assumed to be isobaric, then Qp = ΔH:
ΔH is the enthalpy change of reaction (for a chemical reaction at constant pressure). Then
for a possible process. Let the change ΔG in Gibbs free energy be defined as
(eq.1)
Notice that it is not defined in terms of any external state functions, such as ΔSext or ΔStot. Then the second law becomes, which also tells us about the spontaneity of the reaction:
favoured reaction (Spontaneous)
Neither the forward nor the reverse reaction prevails (Equilibrium)
disfavoured reaction (Nonspontaneous)
Gibbs free energy G itself is defined as
(eq.2)
but notice that to obtain equation (2) from equation (1) we must assume that T is constant. Thus, Gibbs free energy is most useful for thermochemical processes at constant temperature and pressure: both isothermal and isobaric. Such processes don't move on a P-V diagram, such as phase change of a pure substance, which takes place at the saturation pressure and temperature. Chemical reactions, however, do undergo changes in chemical potential, which is a state function. Thus, thermodynamic processes are not confined to the two dimensional P-V diagram. There is a third dimension for n, the quantity of gas. For the study of explosive chemicals, the processes are not necessarily isothermal and isobaric. For these studies, Helmholtz free energy is used.[27]
If an isolated system (Q = 0) is at constant pressure (Q = ΔH), then
Therefore the Gibbs free energy of an isolated system is:
and if ΔG ≤ 0 then this implies that ΔS ≥ 0, back to where we started the derivation of ΔG
The cell is the functional basic unit of life. Cell was discovered by Robert Hooke and is the functional unit of all known living organisms. It is the smallest unit of life that is classified as a living thing, and is often called the building block of life. Some organisms, such as most bacteria, are unicellular (consist of a single cell). Other organisms, such as humans, cats, dogs, and birds, are multicellular. Humans have about 100 trillion or 10 14 cells; a typical cell size is 10 µm and a typical cell mass is 1 nanogram. The largest cells are about 135 µm in the anterior horn in the spinal cord while granule cells in the cerebellum, the smallest, can be some 4 µm and the longest cell can reach from the toe to the lower brain stem (Pseudounipolar cells).
The largest known cells are unfertilised ostrich egg cells which weigh 3.3 pounds.
In 1835, before the final cell theory was developed, Jan Evangelista Purkyně observed small "granules" while looking at the plant tissue through a microscope. The cell theory, first developed in 1839 by Matthias Jakob Schleiden and Theodor Schwann, states that all organisms are composed of one or more cells, that all cells come from preexisting cells, that vital functions of an organism occur within cells, and that all cells contain the hereditary information necessary for regulating cell functions and for transmitting information to the next generation of cells.
The word cell comes from the Latin cellula, meaning, a small room. The descriptive term for the smallest living biological structure was coined by Robert Hooke in a book he published in 1865 when he compared the cork cells he saw through his microscope to the small rooms monks lived in.There are two types of cells: eukaryotic and prokaryotic. Prokaryotic cells are usually independent, while eukaryotic cells are often found in multicellular organisms.[29]
Earth's early atmosphere
Some evidence suggests that Earth's original atmosphere might have contained fewer of the reducing molecules than was thought at the time of the Miller–Urey experiment. There is abundant evidence of major volcanic eruptions 4 billion years ago, which would have released carbon dioxide, nitrogen, hydrogen sulfide (H2S), and sulfur dioxide (SO2) into the atmosphere. Experiments using these gases in addition to the ones in the original Miller–Urey experiment have produced more diverse molecules. The experiment created a mixture that was racemic (containing both L and D enantiomers) and experiments since have shown that "in the lab the two versions are equally likely to appear."[31] However, in nature, L amino acids dominate; later experiments have confirmed disproportionate amounts of L or D oriented enantiomers are possible.[32]
Originally it was thought that the primitive secondary atmosphere contained mostly ammonia and methane. However, it is likely that most of the atmospheric carbon was CO2 with perhaps some CO and the nitrogen mostly N2. In practice gas mixtures containing CO, CO2, N2, etc. give much the same products as those containing CH4 and NH3 so long as there is no O2. The hydrogen atoms come mostly from water vapor. In fact, in order to generate aromatic amino acids under primitive earth conditions it is necessary to use less hydrogen-rich gaseous mixtures. Most of the natural amino acids, hydroxyacids, purines, pyrimidines, and sugars have been made in variants of the Miller experiment.[33]
More recent results may question these conclusions. The University of Waterloo and University of Colorado conducted simulations in 2005 that indicated that the early atmosphere of Earth could have contained up to 40 percent hydrogen—implying a possibly much more hospitable environment for the formation of prebiotic organic molecules. The escape of hydrogen from Earth's atmosphere into space may have occurred at only one percent of the rate previously believed based on revised estimates of the upper atmosphere's temperature.[34] One of the authors, Owen Toon notes: "In this new scenario, organics can be produced efficiently in the early atmosphere, leading us back to the organic-rich soup-in-the-ocean concept... I think this study makes the experiments by Miller and others relevant again." Outgassing calculations using a chondritic model for the early earth complement the Waterloo/Colorado results in re-establishing the importance of the Miller–Urey experiment.[35]
Conditions similar to those of the Miller–Urey experiments are present in other regions of the solar system, often substituting ultraviolet light for lightning as the energy source for chemical reactions. The Murchison meteorite that fell near Murchison, Victoria, Australia in 1969 was found to contain over 90 different amino acids, nineteen of which are found in Earth life. Comets and other icy outer-solar-system bodies are thought to contain large amounts of complex carbon compounds (such as tholins) formed by these processes, darkening surfaces of these bodies.[36] The early Earth was bombarded heavily by comets, possibly providing a large supply of complex organic molecules along with the water and other volatiles they contributed. This has been used to infer an origin of life outside of Earth: the panspermia hypothesis.
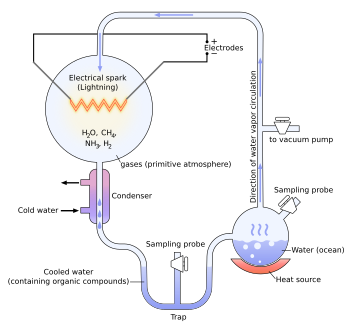
The Miller and Urey experiment[37] (or Urey–Miller experiment)[38] was an experiment that simulated hypothetical conditions thought at the time to be present on the early Earth, and tested for the occurrence of chemical origins of life. Specifically, the experiment tested Alexander Oparin's and J. B. S. Haldane's hypothesis that conditions on the primitive Earth favored chemical reactions that synthesized organic compounds from inorganic precursors. Considered to be the classic experiment on the origin of life, it was conducted in 1952[39] and published in 1953 by Stanley Miller and Harold Urey at the University of Chicago.[40][41][42]
After Miller's death in 2007, scientists examining sealed vials preserved from the original experiments were able to show that there were actually well over 20 different amino acids produced in Miller's original experiments. That is considerably more than what Miller originally reported, and more than the 20 that naturally occur in life. Moreover, some evidence suggests that Earth's original atmosphere might have had a different composition than the gas used in the Miller–Urey experiment. There is abundant evidence of major volcanic eruptions 4 billion years ago, which would have released carbon dioxide, nitrogen, hydrogen sulfide (H2S), and sulfur dioxide (SO2) into the atmosphere. Experiments using these gases in addition to the ones in the original Miller–Urey experiment have produced more diverse molecules.[31]
Experiment
The experiment used water (H2O), methane (CH4), ammonia (NH3), and hydrogen (H2). The chemicals were all sealed inside a sterile array of glass tubes and flasks connected in a loop, with one flask half-full of liquid water and another flask containing a pair of electrodes. The liquid water was heated to induce evaporation, sparks were fired between the electrodes to simulate lightning through the atmosphere and water vapor, and then the atmosphere was cooled again so that the water could condense and trickle back into the first flask in a continuous cycle.
At the end of one week of continuous operation, Miller and Urey observed that as much as 10–15% of the carbon within the system was now in the form of organic compounds. Two percent of the carbon had formed amino acids that are used to make proteins in living cells, with glycine as the most abundant. Sugars, liquids, were also formed. Nucleic acids were not formed within the reaction. But the common 20 amino acids were formed, but in various concentrations.
In an interview, Stanley Miller stated: "Just turning on the spark in a basic pre-biotic experiment will yield 11 out of 20 amino acids."[43]
As observed in all subsequent experiments, both left-handed (L) and right-handed (D) optical isomers were created in a racemic mixture.
One-step reactions among the mixture components can produce hydrogen cyanide (HCN), formaldehyde (CH2O),[45] and other active intermediate compounds (acetylene, cyanoacetylene, etc.):
CO2 → CO + [O] (atomic oxygen)
CH4 + 2[O] → CH2O + H2O
CO + NH3 → HCN + H2O
CH4 + NH3 → HCN + 3H2 (BMA process)
The formaldehyde, ammonia, and HCN then react by Strecker synthesis to form amino acids and other biomolecules:
CH2O + HCN + NH3 → NH2-CH2-CN + H2O
NH2-CH2-CN + 2H2O → NH3 + NH2-CH2-COOH (glycine)
Furthermore, water and formaldehyde can react via Butlerov's reaction to produce various sugars like ribose.
Other experiments
This experiment inspired many others. In 1961, Joan Oró found that the nucleotide base adenine could be made from hydrogen cyanide (HCN) and ammonia in a water solution. His experiment produced a large amount of adenine, which molecules were formed from 5 molecules of HCN.[46]
Also, many amino acids are formed from HCN and ammonia under these conditions.[47]
Experiments conducted later showed that the other RNA and DNA nucleobases could be obtained through simulated prebiotic chemistry with a reducing atmosphere.[48]
There also had been similar electric discharge experiments related to the origin of life contemporaneous with Miller–Urey. An article in The New York Times (March 8, 1953:E9), titled "Looking Back Two Billion Years" describes the work of Wollman (William) M. MacNevin at The Ohio State University, before the Miller Science paper was published in May 1953. MacNevin was passing 100,000 volt sparks through methane and water vapor and produced "resinous solids" that were "too complex for analysis." The article describes other early earth experiments being done by MacNevin. It is not clear if he ever published any of these results in the primary scientific literature.
K. A. Wilde submitted a paper to Science on December 15, 1952, before Miller submitted his paper to the same journal on February 14, 1953. Wilde's paper was published on July 10, 1953.[49] Wilde used voltages up to only 600 V on a binary mixture of carbon dioxide (CO2) and water in a flow system. He observed only small amounts of carbon dioxide reduction to carbon monoxide, and no other significant reduction products or newly formed carbon compounds.
Other researchers were studying UV-photolysis of water vapor with carbon monoxide. They have found that various alcohols, aldehydes and organic acids were synthesized in reaction mixture.[50]
More recent experiments by chemist Jeffrey Bada at Scripps Institution of Oceanography (in La Jolla, CA) were similar to those performed by Miller. However, Bada noted that in current models of early Earth conditions, carbon dioxide and nitrogen (N2) create nitrites, which destroy amino acids as fast as they form. However, the early Earth may have had significant amounts of iron and carbonate minerals able to neutralize the effects of the nitrites. When Bada performed the Miller-type experiment with the addition of iron and carbonate minerals, the products were rich in amino acids. This suggests the origin of significant amounts of amino acids may have occurred on Earth even with an atmosphere containing carbon dioxide and nitrogen.[51].
The sizes of prokaryotes relative to other organisms and biomolecules
Prokaryotes are single-cell organisms that do not have a nucleus, mitochondria, or any other membrane-bound organelles. In other words, neither their DNA nor any of their other sites of metabolic activity are collected together in a discrete membrane-enclosed area. Instead, everything is openly accessible within the cell, some of which is free-floating[3].
A distinction between prokaryotes and eukaryotes (meaning true kernel, also spelled "eucaryotes") is that eukaryotes do have "true" nuclei containing their DNA. Unlike prokaryotes, eukaryotic organisms may be unicellular, as in amoebae, or multicellular, as in plants and animals. The difference between the structure of prokaryotes and eukaryotes is so great that it is sometimes considered to be the most important distinction among groups of organisms.
The cell structure of prokaryotes differs greatly from that of eukaryotes. The defining characteristic is the absence of a nucleus. Also the size of Ribosomes in prokaryotes is smaller than that in eukaryotes, which is now where respiration takes place. The genomes of prokaryotes are held within an irregular DNA/protein complex in the cytosol called the nucleoid, which lacks a nuclear envelope. [52]
In general, prokaryotes lack the following membrane-bound cell compartments: mitochondria and chloroplasts. Instead, processes such as oxidative phosphorylation and photosynthesis take place across the prokaryotic plasma membrane. However, prokaryotes do possess some internal structures, such as cytoskeletons, and the bacterial order Planctomycetes have a membrane around their nucleoid and contain other membrane-bound cellular structures. Both eukaryotes and prokaryotes contain large RNA/protein structures called ribosomes, which produce protein. Prokaryotes are usually much smaller than eukaryotic cells.
Prokaryotes also differ from eukaryotes in that they contain only a single loop of stable chromosomal DNA stored in an area named the nucleoid, whereas eukaryote DNA is found on tightly bound and organized chromosomes. Although some eukaryotes have satellite DNA structures called plasmids, in general these are regarded as a prokaryote feature, and many important genes in prokaryotes are stored on plasmids.
Prokaryotes have a larger surface-area-to-volume ratio giving them a higher metabolic rate, a higher growth rate, and, as a consequence, a shorter generation time compared to Eukaryotes.
A criticism of this classification is that the word "prokaryote" is based on what these organisms are not (they are not eukaryotic), rather than what they are (either archaea or bacteria).
In 1977, Carl Woese proposed dividing prokaryotes into the Bacteria and Archaea (originally Eubacteria and Archaebacteria) because of the major differences in the structure and genetics between the two groups of organisms. This arrangement of Eukaryota (also called "Eukarya"), Bacteria, and Archaea is called the three-domain system, replacing the traditional two-empire system.[53]
The origin of the eukaryotic cell was a milestone in the evolution of life, since they include all complex cells and almost all multi-cellular organisms. The timing of this series of events is hard to determine; Knoll (2006) suggests they developed approximately 1.6 – 2.1 billion years ago. Some acritarchs are known from at least 1,650 million years ago, and the possible alga Grypania has been found as far back as 2,100 million years ago.
Fossils that are clearly related to modern groups start appearing around 1.2 billion years ago, in the form of a red alga, though recent work suggests the existence of fossilized filamentous algae in the Vindhya basin dating back to 1.6 to 1.7 billion years ago.
Biomarkers suggest that at least stem eukaryotes arose even earlier. The presence of steranes in Australian shales indicates that eukaryotes were present 2.7 billion years ago.[54]
There are many different types of eukaryotic cells, though animals and plants are the most familiar eukaryotes, and thus provide an excellent starting point for understanding eukaryotic structure. Fungi and many protists have some substantial differences, however.
Animal cell
An animal cell is a form of eukaryotic cell that makes up many tissues in animals. The animal cell is distinct from other eukaryotes, most notably plant cells, as they lack cell walls and chloroplasts, and they have smaller vacuoles. Due to the lack of a rigid cell wall, animal cells can adopt a variety of shapes, and a phagocytic cell can even engulf other structures.
There are many different cell types. For instance, there are approximately 210 distinct cell types in the adult human body.[55]
Plant cell
Plant cells are quite different from the cells of the other eukaryotic organisms. Their distinctive features are:
A large central vacuole (enclosed by a membrane, the tonoplast), which maintains the cell's turgor and controls movement of molecules between the cytosol and sap
A primary cell wall containing cellulose, hemicellulose and pectin, deposited by the protoplast on the outside of the cell membrane; this contrasts with the cell walls of fungi, which contain chitin, and the cell envelopes of prokaryotes, in which peptidoglycans are the main structural molecules
The plasmodesmata, linking pores in the cell wall that allow each plant cell to communicate with other adjacent cells; this is different from the functionally analogous system of gap junctions between animal cells.
Plastids, especially chloroplasts that contain chlorophyll, the pigment that gives plants their green color and allows them to perform photosynthesis
Higher plants, including conifers and flowering plants (Angiospermae) lack the flagellae and centrioles that are present in animal cells.
Fungal cell
Fungal cells are most similar to animal cells, with the following exceptions:
A cell wall that contains chitin
Less definition between cells; the hyphae of higher fungi have porous partitions called septa, which allow the passage of cytoplasm, organelles, and, sometimes, nuclei. Primitive fungi have few or no septa, so each organism is essentially a giant multinucleate supercell; these fungi are described as coenocytic.
Only the most primitive fungi, chytrids, have flagella.[56]
Other eukaryotic cells
Eukaryotes are a very diverse group, and their cell structures are equally diverse. Many have cell walls; many do not. Many have chloroplasts, derived from primary, secondary, or even tertiary endosymbiosis; and many do not. Some groups have unique structures, such as the cyanelles of the glaucophytes, the haptonema of the haptophytes, or the ejectisomes of the cryptomonads. Other structures, such as pseudopods, are found in various eukaryote groups in different forms, such as the lobose amoebozoans or the reticulose foraminiferans.
Table 1: Comparison of features of Prokaryotic and Eukaryotic cells
Structure of a typical animal cellStructure of a typical plant cell
Plant cells are eukaryotic cells that differ in several key respects from the cells of other eukaryotic organisms. Their distinctive features include: A large central vacuole, a water-filled volume enclosed by a membrane known as the tonoplast maintains the cell's turgor, controls movement of molecules between the cytosol and sap, stores useful material and digests waste proteins and organelles.
A cell wall composed of cellulose and hemicellulose, pectin and in many cases lignin, are secreted by the protoplast on the outside of the cell membrane. This contrasts with the cell walls of fungi (which are made of chitin), and of bacteria, which are made of peptidoglycan.
Specialised cell-cell communication pathways known as plasmodesmata, pores in the primary cell wall through which the plasmalemma and endoplasmic reticulum of adjacent cells are continuous.
Plastids, the notables one being the chloroplasts, which contain chlorophyll and the biochemical systems for light harvesting and photosynthesis, but also amyloplasts specialized for starch storage, elaioplasts specialized for fat storage, and chromoplasts specialized for synthesis and storage of pigments. As in mitochondria, which have a genome encoding 37 genes, plastids have their own genomes of about 100-120 unique genes and, it is presumed, arose as prokaryotic endosymbionts living in the cells of an early eukaryotic ancestor of the land plants and algae.
Unlike animal cells, plant cells are stationary.
Cell division by construction of a phragmoplast as a template for building a cell plate late in cytokinesis is characteristic of land plants and a few groups of algae, the notable one being the Charophytes and the Order Trentepohliales.
The sperm of bryophytes have flagellae similar to those in animals, but higher plants, (including Gymnosperms and flowering plants) lack the flagellae and centrioles that are present in animal cells.[57]
Table 2: Comparison of structures between animal and plant cells
The endosymbiotic (from the Greek: endo- meaning inside and -symbiosis meaning cohabiting) theory was first articulated by the Russian botanist Konstantin Mereschkowski in 1905. Mereschkowski was familiar with work by botanist Andreas Schimper, who had observed in 1883 that the division of chloroplasts in green plants closely resembled that of free-living cyanobacteria, and who had himself tentatively proposed (in a footnote) that green plants had arisen from a symbiotic union of two organisms. Ivan Wallin extended the idea of an endosymbiotic origin to mitochondria in the 1920s.These theories were initially dismissed or ignored. More detailed electron microscopic comparisons between cyanobacteria and chloroplasts (for example studies by Hans Ris), combined with the discovery that plastids and mitochondria contain their own DNA (which by that stage was recognized to be the hereditary material of organisms) led to a resurrection of the idea in the 1960s.
The endosymbiotic theory was advanced and substantiated with microbiological evidence by Lynn Margulis in a 1967 paper, The Origin of Mitosing Eukaryotic Cells.[58]
In her 1981 work Symbiosis in Cell Evolution she argued that eukaryotic cells originated as communities of interacting entities, including endosymbiotic spirochaetes that developed into eukaryotic flagella and cilia. This last idea has not received much acceptance, because flagella lack DNA and do not show ultrastructural similarities to bacteria or archaea. According to Margulis and Dorion Sagan, "Life did not take over the globe by combat, but by networking" (i.e., by cooperation). The possibility that peroxisomes may have an endosymbiotic origin has also been considered, although they lack DNA. Christian de Duve proposed that they may have been the first endosymbionts, allowing cells to withstand growing amounts of free molecular oxygen in the Earth's atmosphere. However, it now appears that they may be formed de novo, contradicting the idea that they have a symbiotic origin.
It is believed that over millennia these endosymbionts transferred some of their own DNA to the host cell's nucleus during the evolutionary transition from a symbiotic community to an instituted eukaryotic cell (called "serial endosymbiosis"). This hypothesis is thought to be possible because it is known today from scientific observation that transfer of DNA occurs between bacteria species, even if they are not closely related. Bacteria can take up DNA from their surroundings and have a limited ability to incorporate it into their own genome.[59]
Eukaryotes are one of the structurally complex cell type, and by definition are in part organized by smaller interior compartments, that are themselves enclosed by lipid membranes that resemble the outermost cell membrane. The larger organelles, such as the nucleus and vacuoles, are easily visible with the light microscope. They were among the first biological discoveries made after the invention of the microscope.[60]
Not all eukaryotic cells have each of the organelles listed below. Exceptional organisms have cells which do not include some organelles that might otherwise be considered universal to eukaryotes (such as mitochondria).[61] There are also occasional exceptions to the number of membranes surrounding organelles, listed in the tables below (e.g., some that are listed as double-membrane are sometimes found with single or triple membranes). In addition, the number of individual organelles of each type found in a given cell varies depending upon the function of that cell.
energy production (house), Mitochondria are self-replicating organelles that occur in various numbers, shapes, and sizes in the cytoplasm of all eukaryotic cells.
double-membrane compartment
most eukaryotes
has some DNA; theorized to be engulfed by an ancestral eukaryotic cell (endosymbiosis)
Mitochondria and chloroplasts, which have double-membranes and their own DNA, are believed to have originated from incompletely consumed or invading prokaryotic organisms, which were adopted as a part of the invaded cell. This idea is supported in the Endosymbiotic theory.
(A) Electron micrograph of Halothiobacillus neapolitanus cells, arrows highlight carboxysomes. (B) Image of intact carboxysomes isolated from H. neapolitanus. Scale bars are 100 nm.[63]
Prokaryotes are not as structurally complex as eukaryotes, and were once thought not to have any internal structures enclosed by lipid membranes. In the past, they were often viewed as having little internal organization; but, slowly, details are emerging about prokaryotic internal structures. An early false turn was the idea developed in the 1970s that bacteria might contain membrane folds termed mesosomes, but these were later shown to be artifacts produced by the chemicals used to prepare the cells for electron microscopy.[64][65]
However, more recent research has revealed that at least some prokaryotes have microcompartments such as carboxysomes. These subcellular compartments are 100 - 200 nm in diameter and are enclosed by a shell of proteins.[66] Even more striking is the description of membrane-bound magnetosomes in bacteria,[67][68] as well as the nucleus-like structures of the Planctomycetes that are surrounded by lipid membranes.[69]
E. coli is frequently used as a model organism in microbiology studies. Cultivated strains (e.g. E. coli K12) are well-adapted to the laboratory environment, and, unlike wild type strains, have lost their ability to thrive in the intestine. Many lab strains lose their ability to form biofilms. These features protect wild type strains from antibodies and other chemical attacks, but require a large expenditure of energy and material resources.
In 1946, Joshua Lederberg and Edward Tatum first described the phenomenon known as bacterial conjugation using E. coli as a model bacterium, and it remains the primary model to study conjugation.[citation needed] E. coli was an integral part of the first experiments to understand phage genetics, and early researchers, such as Seymour Benzer, used E. coli and phage T4 to understand the topography of gene structure. Prior to Benzer's research, it was not known whether the gene was a linear structure, or if it had a branching pattern.
E. coli was one of the first organisms to have its genome sequenced; the complete genome of E. coli K12 was published by Science in 1997.
The long-term evolution experiments using E. coli, begun by Richard Lenski in 1988, have allowed direct observation of major evolutionary shifts in the laboratory. In this experiment, one population of E. coli unexpectedly evolved the ability to aerobically metabolize citrate. This capacity is extremely rare in E. coli. As the inability to grow aerobically is normally used as a diagnostic criterion with which to differentiate E. coli from other, closely related bacteria such as Salmonella, this innovation may mark a speciation event observed in the lab.
By combining nanotechnologies with landscape ecology complex habitat landscapes can be generated with details at the nanoscale. On such synthetic ecosystems evolutionary experiments with E. coli have been performed in order to study the spatial biophysics of adaptation in an island biogeography on-chip.
Because of its long history of laboratory culture and ease of manipulation, E. coli also plays an important role in modern biological engineering and industrial microbiology. The work of Stanley Norman Cohen and Herbert Boyer in E. coli, using plasmids and restriction enzymes to create recombinant DNA, became a foundation of biotechnology.
Considered a very versatile host for the production of heterologous proteins, researchers can introduce genes into the microbes using plasmids, allowing for the mass production of proteins in industrial fermentation processes. Genetic systems have also been developed which allow the production of recombinant proteins using E. coli. One of the first useful applications of recombinant DNA technology was the manipulation of E. coli to produce human insulin. Modified E. coli have been used in vaccine development, bioremediation, and production of immobilised enzymes. E. coli cannot, however, be used to produce some of the more large, complex proteins which contain multiple disulfide bonds and, in particular, unpaired thiols, or proteins that also require post-translational modification for activity.
Studies are also being performed into programming E. coli to potentially solve complicated mathematics problems such as the Hamiltonian path problem.
The yeast cell's life cycle: 1. Budding 2. Conjugation 3. Spore
A model organism
When researchers look for an organism to use in their studies, they look for several traits. Among these are size, generation time, accessibility, manipulation, genetics, conservation of mechanisms, and potential economic benefit. The yeast species S. pombe and S. cerevisiae are both well studied; these two species diverged approximately 300 to 600 million years before present, and are significant tools in the study of DNA damage and repair mechanisms.
The alpha-factor of S. cerevisiae, has been compared to the liphophilic peptide created by the fungus Tremella mesenterica.
S. cerevisiae has developed as a model organism because it scores favorably on a number of these criteria.
As a single celled organism S. cerevisiae is small with a short generation time (doubling time 1.25–2 hours at 30 °C (86 °F)) and can be easily cultured. These are all positive characteristics in that they allow for the swift production and maintenance of multiple specimen lines at low cost.
S. cerevisiae can be transformed allowing for either the addition of new genes or deletion through homologous recombination. Furthermore, the ability to grow S. cerevisiae as a haploid simplifies the creation of gene knockouts strains.
As a eukaryote, S. cerevisiae shares the complex internal cell structure of plants and animals without the high percentage of non-coding DNA that can confound research in higher eukaryotes.
S. cerevisiae research is a strong economic driver, at least initially, as a result of its established use in industry.[70]
Genome sequencingS. cerevisiae was the first eukaryotic genome that was completely sequenced. The genome sequence was released in the public domain on April 24, 1996. Since then, regular updates have been maintained at the Saccharomyces Genome Database (SGD). This database is a highly annotated and cross-referenced database for yeast researchers. Another important S. cerevisiae database is maintained by the Munich Information Center for Protein Sequences (MIPS). The genome is composed of about 12,156,677 base pairs and 6,275 genes, compactly organized on 16 chromosomes. Only about 5,800 of these are believed to be true functional genes. Yeast is estimated to share about 23% of its genome with that of humans.
The availability of the S. cerevisiae genome sequence and the complete set of deletion mutants has further enhanced the power of S. cerevisiae as a model for understanding the regulation of eukaryotic cells. A project underway to analyze the genetic interactions of all double deletion mutants through synthetic genetic array analysis will take this research one step further.
Approaches that can be applied in many different fields of biological and medicinal science have been developed by yeast scientists. These include yeast two-hybrid for studying protein interactions and tetrad analysis.
↑Dubos J. (1951). "Louis Pasteur: Free Lance of Science, Gollancz. Quoted in Manchester K. L. (1995) Louis Pasteur (1822–1895)--chance and the prepared mind". Trends Biotechnol. 13 (12): 511–515. doi:10.1016/S0167-7799(00)89014-9. PMID8595136.
↑Kornberg H, Krebs H (1957). "Synthesis of cell constituents from C2-units by a modified tricarboxylic acid cycle". Nature. 179 (4568): 988–91. doi:10.1038/179988a0. PMID13430766.
↑Robert K. Crane, D. Miller and I. Bihler. “The restrictions on possible mechanisms of intestinal transport of sugars”. In: Membrane Transport and Metabolism. Proceedings of a Symposium held in Prague, 22–27 August 1960. Edited by A. Kleinzeller and A. Kotyk. Czech Academy of Sciences, Prague, 1961, pp. 439-449.
↑Thompson WR, Murray BG, Khare BN, Sagan C (1987). "Coloration and darkening of methane clathrate and other ices by charged particle irradiation: applications to the outer solar system". Journal of geophysical research. 92 (A13): 14933–47. doi:10.1029/JA092iA13p14933. PMID11542127. {{cite journal}}: Unknown parameter |month= ignored (help)CS1 maint: multiple names: authors list (link)
↑Hill HG, Nuth JA (2003). "The catalytic potential of cosmic dust: implications for prebiotic chemistry in the solar nebula and other protoplanetary systems". Astrobiology. 3 (2): 291–304. doi:10.1089/153110703769016389. PMID14577878.
↑Balm SP, Hare J.P., Kroto HW (1991). "The analysis of comet mass spectrometric data". Space Science Reviews. 56: 185–9. doi:10.1007/BF00178408.{{cite journal}}: CS1 maint: multiple names: authors list (link)
↑Miller, Stanley L. (1959). "Organic Compound Synthesis on the Primitive Earth". Science. 130 (3370): 245. doi:10.1126/science.130.3370.245. PMID13668555. {{cite journal}}: Unknown parameter |coauthors= ignored (|author= suggested) (help); Unknown parameter |month= ignored (help) Miller states that he made "A more complete analysis of the products" in the 1953 experiment, listing additional results.
↑A. Lazcano, J. L. Bada (2004). "The 1953 Stanley L. Miller Experiment: Fifty Years of Prebiotic Organic Chemistry". Origins of Life and Evolution of Biospheres. 33 (3): 235–242. doi:10.1023/A:1024807125069. PMID14515862. {{cite journal}}: Unknown parameter |month= ignored (help)
↑Oró J, Kimball AP (1961). "Synthesis of purines under possible primitive earth conditions. I. Adenine from hydrogen cyanide". Archives of biochemistry and biophysics. 94: 217–27. doi:10.1016/0003-9861(61)90033-9. PMID13731263. {{cite journal}}: Unknown parameter |month= ignored (help)
↑Oró J, Kamat SS (1961). "Amino-acid synthesis from hydrogen cyanide under possible primitive earth conditions". Nature. 190: 442–3. doi:10.1038/190442a0. PMID13731262. {{cite journal}}: Unknown parameter |month= ignored (help)
↑Oró J (1967). Fox SW (ed.). Origins of Prebiological Systems and of Their Molecular Matrices. New York Academic Press. p. 137.
↑Ryter A (1988). "Contribution of new cryomethods to a better knowledge of bacterial anatomy". Ann. Inst. Pasteur Microbiol. 139 (1): 33–44. doi:10.1016/0769-2609(88)90095-6. PMID3289587.
↑Kerfeld CA, Sawaya MR, Tanaka S, Nguyen CV, Phillips M, Beeby M, Yeates TO (2005). "Protein structures forming the shell of primitive bacterial organelles". Science. 309 (5736): 936–8. doi:10.1126/science.1113397. PMID16081736. {{cite journal}}: Unknown parameter |month= ignored (help)CS1 maint: multiple names: authors list (link)
↑Komeili A, Li Z, Newman DK, Jensen GJ (2006). "Magnetosomes are cell membrane invaginations organized by the actin-like protein MamK". Science. 311 (5758): 242–5. doi:10.1126/science.1123231. PMID16373532.{{cite journal}}: CS1 maint: multiple names: authors list (link)
↑Scheffel A, Gruska M, Faivre D, Linaroudis A, Plitzko JM, Schüler D (2006). "An acidic protein aligns magnetosomes along a filamentous structure in magnetotactic bacteria". Nature. 440 (7080): 110–4. doi:10.1038/nature04382. PMID16299495.{{cite journal}}: CS1 maint: multiple names: authors list (link)