The term Molecular biology was first used by Warren Weaver in 1938. Molecular biology is the study of molecular underpinnings of the processes of replication, transcription, translation, and cellular function.
A typical animal cell. Within the cytoplasm, the major organelles and cellular structures include: (1) nucleolus (2) nucleus (3) ribosome (4) vesicle (5) rough endoplasmic reticulum (6) Golgi apparatus (7) cytoskeleton (8) smooth endoplasmic reticulum (9) mitochondria (10) vacuole (11) cytosol (12) lysosome (13) centriole.
The term macromolecule was coined by Nobel laureate Hermann Staudinger in the 1920s, although his first relevant publication on this field only mentioned high molecular compounds (in excess of 1000 atoms). At that time the phrase polymer as introduced by Berzelius in 1833 had a different meaning from that of today: it simply was another form of isomerism, such as an enzene or acetylene, and had little to do with size. Some examples of organic macromolecules are bio-polymers (carbohydrates, proteins, lipids, nucleic acids) or polymers (plastics, synthetic fiber and rubber).
A carbohydrate (kɑ:bəˈhaɪdreɪt/) is an organic compound which has the empirical formula Cm(H2O)n; that is, consists only of carbon, hydrogen and oxygen, with a hydrogen:oxygen atom ratio of 2:1 (as in water). Carbohydrates can be viewed as hydrates of carbon, hence their name. Structurally however, it is more accurate to view them as polyhydroxy aldehydes and ketones. Historically nutritionists have classified carbohydrates as either simple or complex, however, the exact delineation of these categories is ambiguous. Today, the term simple carbohydrate typically refers to monosaccharides and disaccharides, and complex carbohydrate means polysaccharides (and oligosaccharides).
Monosaccharides (from Greek monos: single, sacchar: sugar) are the most basic units of biologically important carbohydrates. They are the simplest form of sugar and are usually colorless, water-soluble, crystalline solids. Some monosaccharides have a sweet taste. Examples of monosaccharides include glucose (dextrose), fructose (levulose), galactose, xylose and ribose. Monosaccharides are the building blocks of disaccharides such as sucrose and polysaccharides (such as cellulose and starch). Further, each carbon atom that supports a hydroxyl group (except for the first and last) is chiral, giving rise to a number of isomeric forms all with the same chemical formula. For instance, galactose and glucose are both aldohexoses, but have different chemical and physical properties.
A disaccharide or biose is the carbohydrate formed when two monosaccharides undergo a condensation reaction which involves the elimination of a small molecule, such as water, from the functional groups only. Like monosaccharides, disaccharides also dissolve in water, taste sweet and are called sugars. The glycosidic bond can be formed between any hydroxyl group on the component monosaccharide. So, even if both component sugars are the same (e.g., glucose), different bond combinations (regiochemistry) and stereochemistry (alpha- or beta-) result in disaccharides that are diastereoisomers with different chemical and physical properties.
Depending on the monosaccharide constituents, disaccharides are sometimes crystalline, sometimes water-soluble, and sometimes sweet-tasting and sticky-feeling.
Disaccharide
Unit 1
Unit 2
Bond
Sucrose (table sugar, cane sugar, beet sugar, or saccharose)
An oligosaccharide (from the Greek oligos, a few, and sacchar, sugar) is a saccharide polymer containing typically three to ten component sugars, also known as many as 8 sugars, or polysaccharides. Oligosaccharides can have many functions; for example, they are commonly found on the plasma membrane of animal cells where they can play a role in cell-cell recognition. In general, they are found either O- or N-linked to compatible amino acid side-chains in proteins or to lipid moieties.
e.g.Fructo-oligosaccharides (FOS), which are found in many vegetables, consist of short chains of fructose molecules. (Inulin has a much higher degree of polymerization than FOS and is a polysaccharide.) Galactooligosaccharides (GOS), which also occur naturally, consist of short chains of galactose molecules. These compounds can be only partially digested by humans.
Polysaccharides are polymeric carbohydrate structures, formed of repeating units (either mono- or di-saccharides) joined together by glycosidic bonds. These structures are often linear, but may contain various degrees of branching. Polysaccharides are often quite heterogeneous, containing slight modifications of the repeating unit. Depending on the structure, these macromolecules can have distinct properties from their monosaccharide building blocks. They may be amorphous or even insoluble in water.Starches are glucose polymers in which glucopyranose units are bonded by alpha-linkages. It is made up of a mixture of Amylose (15–20%) and Amylopectin (80–85%). Amylose consists of a linear chain of several hundred glucose molecules and Amylopectin is a branched molecule made of several thousand glucose units (every chain 24–30 glucose unit). Starches are insoluble in water. They can be digested by hydrolysis, catalyzed by enzymes called amylases, which can break the alpha-linkages (glycosidic bonds). Humans and other animals have amylases, so they can digest starches. Potato, rice, wheat, and maize are major sources of starch in the human diet. The formation of starches are the way that plants store glucose.Glycogen is a polysaccharide that is found in animals and is composed of a branched chain of glucose residues. It is stored in liver and muscles.Chitin is one of many naturally occurring polymers. It is one of the most abundant natural materials in the world. Over time it is bio-degradable in the natural environment. Its breakdown may be catalyzed by enzymes called chitinases, secreted by microorganisms such as bacteria and fungi, and produced by some plants.Arabinoxylans are the copolymers of two pentose sugars - arabinose and xylose.
Proteins are polymer of amino acids linked together by peptide bonds. Amino acids can be divided into two group essential amino acids and non-essential amino acids. Proteins and carbohydrates contain 4 kcal/gram as opposed to lipids which contain 9 kcal/ gram. The liver, and to a much lesser extent the kidneys, can convert amino acids used by cells in protein biosynthesis into glucose by a process known as gluconeogenesis.
The essential amino acids, which must be obtained from external sources (food), are leucine, isoleucine, valine, lysine, threonine, tryptophan, methionine, phenylalanine and histidine. On the other hand, non-essential amino acids are synthesized in our body from other amino acids. The non-essential amino acids are arginine, alanine, asparagine, aspartic acid, cysteine, glutamine, glutamic acid, glycine, proline, serine, and tyrosine.
Proteins( /ˈproʊtiːnz/; also known as polypeptides) are organic compounds made of amino acids arranged in a linear chain and folded into a globular or fibrous form. The amino acids in a polymer are joined together by the peptide bonds between the carboxyl and amino groups of adjacent amino acid residues. The sequence of amino acids in a protein is defined by the sequence of a gene, which is encoded in the genetic code.Proteins were first described by the Dutch chemist Gerhardus Johannes Mulder and named by the Swedish chemist Jöns Jakob Berzelius in 1838.
Lipids are a broad group of naturally occurring molecules which includes fats, waxes, sterols, fat-soluble vitamins (such as vitamins A, D, E and K), monoglycerides, diglycerides, phospholipids, and others. The main biological functions of lipids include energy storage, as structural components of cell membranes, and as important signaling molecules. Cell contain near about 70% of water.
Lipids in membrane
Eukaryotic cells are compartmentalized into membrane-bound organelles which carry out different biological functions. The glycerophospholipids are the main structural component of biological membranes, such as the cellular plasma membrane and the intracellular membranes of organelles; in animal cells the plasma membrane physically separates the intracellular components from the extracellular environment. The glycerophospholipids are amphipathic molecules (containing both hydrophobic and hydrophilic regions) that contain a glycerol core linked to three fatty acid-derived "tails" by ester linkages and to one "head" group by a phosphate ester linkage. While glycerophospholipids are the major component of biological membranes, other non-glyceride lipid components such as sphingomyelin and sterols (mainly cholesterol in animal cell membranes) are also found in biological membranes. In plants and algae, the galactosyldiacylglycerols, and sulfoquinovosyldiacylglycerol, which lack a phosphate group, are important components of membranes of chloroplasts and related organelles and are the most abundant lipids in photosynthetic tissues, including those of higher plants, algae and certain bacteria.
Bilayers have been found to exhibit high levels of birefringence which can be used to probe the degree of order (or disruption) within the bilayer using techniques such as dual polarisation interferometry.
A biological membrane is a form of lipid bilayer. The formation of lipid bilayers is an energetically preferred process when the glycerophospholipids described above are in an aqueous environment. In an aqueous system, the polar heads of lipids align towards the polar, aqueous environment, while the hydrophobic tails minimize their contact with water and tend to cluster together, forming a vesicle; depending on the concentration of the lipid, this biophysical interaction may result in the formation of micelles, liposomes, or lipid bilayers. Other aggregations are also observed and form part of the polymorphism of amphiphile (lipid) behavior. Phase behavior is an area of study within biophysics and is the subject of current academic research. Micelles and bilayers form in the polar medium by a process known as the hydrophobic effect. When dissolving a lipophilic or amphiphilic substance in a polar environment, the polar molecules (i.e., water in an aqueous solution) become more ordered around the dissolved lipophilic substance, since the polar molecules cannot form hydrogen bonds to the lipophilic areas of the amphiphile. So in an aqueous environment, the water molecules form an ordered "clathrate" cage around the dissolved lipophilic molecule.
Role of lipid in signalling
In recent years, evidence has emerged showing that lipid signaling is a vital part of the cell signaling. Lipid signaling may occur via activation of G protein-coupled or nuclear receptors, and members of several different lipid categories have been identified as signaling molecules and cellular messengers. These include sphingosine-1-phosphate, a sphingolipid derived from ceramide that is a potent messenger molecule involved in regulating calcium mobilization, cell growth, and apoptosis; diacylglycerol (DAG) and the phosphatidylinositol phosphates (PIPs), involved in calcium-mediated activation of protein kinase C; the prostaglandins, which are one type of fatty-acid derived eicosanoid involved in inflammation and immunity; the steroid hormones such as estrogen, testosterone and cortisol, which modulate a host of functions such as reproduction, metabolism and blood pressure; and the oxysterols such as 25-hydroxy-cholesterol that are liver X receptor agonists.
We know water is essential to all life as we know it so much so that every cell is nearly 70% of water. We also know that one molecule of water is made up of two hydrogen (H) atoms which are covalently bonded to a single oxygen(O) atom (water's chemical formula H2O).
A noncovalent bond is a type of chemical bond, typically between macromolecules, that does not involve the sharing of pairs of electrons, but rather involves more dispersed variations of electromagnetic interactions. The noncovalent bond is the dominant type of bond between supermolecules in supermolecular chemistry. Noncovalent bonds are critical in maintaining the three-dimensional structure of large molecules, such as proteins and nucleic acids, and are involved in many biological processes in which large molecules bind specifically but transiently to one another. The energy released in the formation of noncovalent bonds is on the order of 1-5 kcal per mol. There are four commonly mentioned types of non-covalent interactions: hydrogen bonds, ionic bonds, van der Waals forces, and hydrophobic interactions. The noncovalent interactions hold together the two strands of DNA in the double helix, stabilize secondary and tertiary structures of proteins, and enable enzyme-substrate binding and antibody-antigen association.
Intramolecular noncovalent interactions are largely responsible for the secondary and tertiary structure of proteins and therefore the protein's function in the mechanisms of life. Intermolecular noncovalent interactions are responsible for protein complexes (quaternary structure) where two or more proteins function in a coherent mechanism.
Most drugs work by noncovalently interacting with biomolecules such as proteins or RNA. Relatively few drugs actually form covalent bonds with the biomolecules they interact with; instead, they interfere with or activate some biological mechanism through noncovalently interacting in very specific locations on specific biomolecules which present the perfect combination of noncovalent binding partners in just the right geometry.
Hydrogen bonding
The best example of a hydrogen bond is found between water molecules.Water molecules contains two hydrogen atoms and one oxygen atom.Two molecules of water can form a hydrogen bond between them. Inside the cell hydrogen bonding also plays an important role in determining the 3D structures of proteins and nucleic bases. In these macromolecules, bonding between parts of the same macromolecule cause it to fold into a specific shape, which helps determine the molecule's physiological or biochemical function. The double helical structure of DNA, for example, is due to hydrogen bonding between the base pairs, which link one complementary strand to the other and enable replication.Hydrogen bonds are also important in the structure of macromolecule cellulose. We should also remember that the hydrogen bond is stronger than a van der Waals interaction, but generally weaker than ionic bonds.
As we know very well in chemistry, pH is a measure of the acidity or basicity of an aqueous solution. Pure water is said to be neutral, with a pH close to 7.0 at 25 °C (77 °F). Solutions with a pH less than 7 are said to be acidic and solutions with a pH greater than 7 are basic or alkaline. pH measurements are important in medicine, biology, chemistry, food science, environmental science, oceanography, civil engineering and many other applications.
In a solution pH approximates but is not equal to p[H], the negative logarithm (base 10) of the molar concentration of dissolved hydronium ions (H3O+); a low pH indicates a high concentration of hydronium ions, while a high pH indicates a low concentration. Crudely, this negative of the logarithm matches the number of places behind the decimal point, so for example 0.1 molar hydrochloric acid should be near pH 1 and 0.0001 molar HCl should be near pH 4 (the base 10 logarithms of 0.1 and 0.0001 being −1, and −4, respectively). Pure (de-ionised) water is neutral, and can be considered either a very weak acid or a very weak base (center of the 0 to 14 pH scale), giving it a pH of 7 (at 25 °C (77 °F)), or 0.0000001 M H+. For an aqueous solution to have a higher pH, a base must be dissolved in it, which binds away many of these rare hydrogen ions. Hydrogen ions in water can be written simply as H+ or as hydronium (H3O+) or higher species (e.g. H9O4+) to account for solvation, but all describe the same entity.
Most of the Earth's freshwater surface bodies are slightly acidic due to the abundance and absorption of carbon dioxide; in fact, for millennia in the past most fresh water bodies have long existed at a slightly acidic pH level.
However, pH is not precisely p[H], but takes into account an activity factor. This represents the tendency of hydrogen ions to interact with other components of the solution, which affects among other things the electrical potential read using a pH meter. As a result, pH can be affected by the ionic strength of a solution – for example, the pH of a 0.05 M potassium hydrogen phthalate solution can vary by as much as 0.5 pH units as a function of added potassium chloride, even though the added salt is neither acidic nor basic.pH also play important role in living organism.
The pH of different cellular compartments, body fluids, and organs is usually tightly regulated in a process called acid-base homeostasis.
The pH of blood is usually slightly basic with a value of pH 7.365. This value is often referred to as physiological pH in biology and medicine.
Plaque can create a local acidic environment that can result in tooth decay by demineralisation.
Enzymes and other proteins have an optimum pH range and can become inactivated or denatured outside this range.
The most common disorder in acid-base homeostasis is acidosis, a condition in which there is an acid overload in the body, generally defined by pH falling below 7.35.
In the blood, pH can be estimated from known base excess (be) and bicarbonate concentration (HCO3) by the following equation:
The cell is the functional basic unit of life. It was discovered by Robert Hooke and is the functional unit of all known living organisms. It is the smallest unit of life that is classified as a living thing, and is often called the building block of life. Some organisms, such as most bacteria, are unicellular (consist of a single cell). Other organisms, such as humans and birds, are multicellular. Humans have about 100 trillion or 1014 cells; a typical cell size is 10 µm and a typical cell mass is 1 nanogram. The largest cells are about 135 µm in the anterior horn in the spinal cord while granule cells in the cerebellum, the smallest, can be some 4 µm and the longest cell can reach from the toe to the lower brain stem (Pseudounipolar cells).
The largest known cells are unfertilised ostrich egg cells which weigh 3.3 pounds.
In 1835, before the final cell theory was developed, Jan Evangelista Purkyně observed small "granules" while looking at the plant tissue through a microscope. The cell theory, first developed in 1839 by Matthias Jakob Schleiden and Theodor Schwann, states that all organisms are composed of one or more cells, that all cells come from preexisting cells, that vital functions of an organism occur within cells, and that all cells contain the hereditary information necessary for regulating cell functions and for transmitting information to the next generation of cells.
The word cell comes from the Latin cellula, meaning, a small room. The descriptive term for the smallest living biological structure was coined by Robert Hooke in a book he published in 1665 when he compared the cork cells he saw through his microscope to the small rooms monks lived in.There are two types of cells: eukaryotic and prokaryotic. Prokaryotic cells are usually independent, while eukaryotic cells are often found in multicellular organisms.
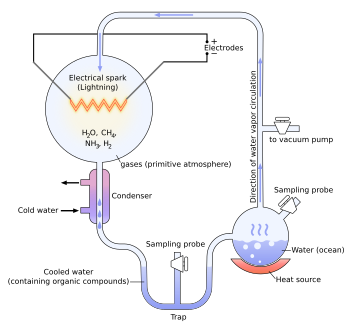
Origin of life and Miller's experiment
The experiment
Earth's early atmosphere
Some evidence suggests that Earth's original atmosphere might have contained fewer of the reducing molecules than was thought at the time of the Miller–Urey experiment. There is abundant evidence of major volcanic eruptions 4 billion years ago, which would have released carbon dioxide, nitrogen, hydrogen sulfide (H2S), and sulfur dioxide (SO2) into the atmosphere. Experiments using these gases in addition to the ones in the original Miller–Urey experiment have produced more diverse molecules. The experiment created a mixture that was racemic (containing both L and D enantiomers) and experiments since have shown that "in the lab the two versions are equally likely to appear."[1] However, in nature, L amino acids dominate; later experiments have confirmed disproportionate amounts of L or D oriented enantiomers are possible.[2]
Originally it was thought that the primitive secondary atmosphere contained mostly ammonia and methane. However, it is likely that most of the atmospheric carbon was CO2 with perhaps some CO and the nitrogen mostly N2. In practice gas mixtures containing CO, CO2, N2, etc. give much the same products as those containing CH4 and NH3 so long as there is no O2. The hydrogen atoms come mostly from water vapor. In fact, in order to generate aromatic amino acids under primitive earth conditions it is necessary to use less hydrogen-rich gaseous mixtures. Most of the natural amino acids, hydroxyacids, purines, pyrimidines, and sugars have been made in variants of the Miller experiment.[3]
More recent results may question these conclusions. The University of Waterloo and University of Colorado conducted simulations in 2005 that indicated that the early atmosphere of Earth could have contained up to 40 percent hydrogen—implying a possibly much more hospitable environment for the formation of prebiotic organic molecules. The escape of hydrogen from Earth's atmosphere into space may have occurred at only one percent of the rate previously believed based on revised estimates of the upper atmosphere's temperature.[4] One of the authors, Owen Toon notes: "In this new scenario, organics can be produced efficiently in the early atmosphere, leading us back to the organic-rich soup-in-the-ocean concept... I think this study makes the experiments by Miller and others relevant again." Outgassing calculations using a chondritic model for the early earth complement the Waterloo/Colorado results in re-establishing the importance of the Miller–Urey experiment.[5]
Conditions similar to those of the Miller–Urey experiments are present in other regions of the solar system, often substituting ultraviolet light for lightning as the energy source for chemical reactions. The Murchison meteorite that fell near Murchison, Victoria, Australia in 1969 was found to contain over 90 different amino acids, nineteen of which are found in Earth life. Comets and other icy outer-solar-system bodies are thought to contain large amounts of complex carbon compounds (such as tholins) formed by these processes, darkening surfaces of these bodies.[6] The early Earth was bombarded heavily by comets, possibly providing a large supply of complex organic molecules along with the water and other volatiles they contributed. This has been used to infer an origin of life outside of Earth: the panspermia hypothesis.
The Miller and Urey experiment[7] (or Urey–Miller experiment)[8] was an experiment that simulated hypothetical conditions thought at the time to be present on the early Earth, and tested for the occurrence of chemical origins of life. Specifically, the experiment tested Alexander Oparin's and J. B. S. Haldane's hypothesis that conditions on the primitive Earth favored chemical reactions that synthesized [organic compounds from inorganic precursors. Considered to be the classic experiment on the origin of life, it was conducted in 1952[9] and published in 1953 by Stanley Miller and Harold Urey at the University of Chicago.[10][11][12]
After Miller's death in 2007, scientists examining sealed vials preserved from the original experiments were able to show that there were actually well over 20 different amino acids produced in Miller's original experiments. That is considerably more than what Miller originally reported, and more than the 20 that naturally occur in life. Moreover, some evidence suggests that Earth's original atmosphere might have had a different composition than the gas used in the Miller–Urey experiment. There is abundant evidence of major volcanic eruptions 4 billion years ago, which would have released carbon dioxide, nitrogen, hydrogen sulfide (H2S), and sulfur dioxide (SO2) into the atmosphere. Experiments using these gases in addition to the ones in the original Miller–Urey experiment have produced more diverse molecules.[1]
Experiment
The experiment used water (H2O), methane (CH4), ammonia (NH3), and hydrogen (H2). The chemicals were all sealed inside a sterile array of glass tubes and flasks connected in a loop, with one flask half-full of liquid water and another flask containing a pair of electrodes. The liquid water was heated to induce evaporation, sparks were fired between the electrodes to simulate lightning through the atmosphere and water vapor, and then the atmosphere was cooled again so that the water could condense and trickle back into the first flask in a continuous cycle.
At the end of one week of continuous operation, Miller and Urey observed that as much as 10–15% of the carbon within the system was now in the form of organic compounds. Two percent of the carbon had formed amino acids that are used to make proteins in living cells, with glycine as the most abundant. Sugars, liquids, were also formed. Nucleic acids were not formed within the reaction. But the common 20 amino acids were formed, but in various concentrations.
In an interview, Stanley Miller stated: "Just turning on the spark in a basic pre-biotic experiment will yield 11 out of 20 amino acids."[13]
As observed in all subsequent experiments, both left-handed (L) and right-handed (D) optical isomers were created in a racemic mixture.
The original experiment remains today under the care of Miller and Urey's former student Professor Jeffrey Bada at the University of California, San Diego, Scripps Institution of Oceanography.[14]
One-step reactions among the mixture components can produce hydrogen cyanide (HCN), formaldehyde (CH2O),[15] and other active intermediate compounds (acetylene, cyanoacetylene, etc.):
CO2 → CO + [O] (atomic oxygen)
CH4 + 2[O] → CH2O + H2O
CO + NH3 → HCN + H2O
CH4 + NH3 → HCN + 3H2 (BMA process)
The formaldehyde, ammonia, and HCN then react by Strecker synthesis to form amino acids and other biomolecules:
Furthermore, water and formaldehyde can react via Butlerov's reaction to produce various sugars like ribose.
Other experiments
This experiment inspired many others. In 1961, Joan Oró found that the nucleotide base adenine could be made from hydrogen cyanide (HCN) and ammonia in a water solution. His experiment produced a large amount of adenine, which molecules were formed from 5 molecules of HCN.[16]
Also, many amino acids are formed from HCN and ammonia under these conditions.[17]
Experiments conducted later showed that the other RNA and DNA nucleobases could be obtained through simulated prebiotic chemistry with a reducing atmosphere.[18]
There also had been similar electric discharge experiments related to the origin of life contemporaneous with Miller–Urey. An article in The New York Times (March 8, 1953:E9), titled "Looking Back Two Billion Years" describes the work of Wollman (William) M. MacNevin at The Ohio State University, before the Miller Science paper was published in May 1953. MacNevin was passing 100,000 volt sparks through methane and water vapor and produced "resinous solids" that were "too complex for analysis." The article describes other early earth experiments being done by MacNevin. It is not clear if he ever published any of these results in the primary scientific literature.[citation needed]
K. A. Wilde submitted a paper to Science on December 15, 1952, before Miller submitted his paper to the same journal on February 14, 1953. Wilde's paper was published on July 10, 1953.[19] Wilde used voltages up to only 600 V on a binary mixture of carbon dioxide (CO2) and water in a flow system. He observed only small amounts of carbon dioxide reduction to carbon monoxide, and no other significant reduction products or newly formed carbon compounds.
Other researchers were studying UV-photolysis of water vapor with carbon monoxide. They have found that various alcohols, aldehydes and organic acids were synthesized in reaction mixture.[20]
More recent experiments by chemist Jeffrey Bada atScripps Institution of Oceanography (in La Jolla, CA) were similar to those performed by Miller. However, Bada noted that in current models of early Earth conditions, carbon dioxide and nitrogen (N2) create nitrite]s, which destroy amino acids as fast as they form. However, the early Earth may have had significant amounts of iron and carbonate minerals able to neutralize the effects of the nitrites. When Bada performed the Miller-type experiment with the addition of iron and carbonate minerals, the products were rich in amino acids. This suggests the origin of significant amounts of amino acids may have occurred on Earth even with an atmosphere containing carbon dioxide and nitrogen.[21]
The sizes of prokaryotes relative to other organisms and biomolecules
Prokaryotes are single-cell organisms that do not have a nucleus, mitochondria, or any other membrane-bound organelles. In other words, neither their DNA nor any of their other sites of metabolic activity are collected together in a discrete membrane-enclosed area. Instead, everything is openly accessible within the cell, some of which is free-floating.
A distinction between prokaryotes and eukaryotes (meaning true kernel, also spelled "eucaryotes") is that eukaryotes do have "true" nuclei containing their DNA. Unlike prokaryotes, eukaryotic organisms may be unicellular, as in amoebae, or multicellular, as in plants and animals. The difference between the structure of prokaryotes and eukaryotes is so great that it is sometimes considered to be the most important distinction among groups of organisms.
The cell structure of prokaryotes differs greatly from that of eukaryotes. The defining characteristic is the absence of a nucleus. Also the size of Ribosomes in prokaryotes is smaller than that in eukaryotes, which is now where respiration takes place. The genomes of prokaryotes are held within an irregular DNA/protein complex in the cytosol called the nucleoid, which lacks a nuclear envelope.
In general, prokaryotes lack the following membrane-bound cell compartments: mitochondria and chloroplasts. Instead, processes such as oxidative phosphorylation and photosynthesis take place across the prokaryotic plasma membrane. However, prokaryotes do possess some internal structures, such as cytoskeletons, and the bacterial order Planctomycetes have a membrane around their nucleoid and contain other membrane-bound cellular structures. Both eukaryotes and prokaryotes contain large RNA/protein structures called ribosomes, which produce protein. Prokaryotes are usually much smaller than eukaryotic cells.
Prokaryotes also differ from eukaryotes in that they contain only a single loop of stable chromosomal DNA stored in an area named the nucleoid, whereas eukaryote DNA is found on tightly bound and organized chromosomes. Although some eukaryotes have satellite DNA structures called plasmids, in general these are regarded as a prokaryote feature, and many important genes in prokaryotes are stored on plasmids.
Prokaryotes have a larger surface-area-to-volume ratio giving them a higher metabolic rate, a higher growth rate, and, as a consequence, a shorter generation time compared to Eukaryotes.
A criticism of this classification is that the word "prokaryote" is based on what these organisms are not (they are not eukaryotic), rather than what they are (either archaea or bacteria).
In 1977, Carl Woese proposed dividing prokaryotes into the Bacteria and Archaea (originally Eubacteria and Archaebacteria) because of the major differences in the structure and genetics between the two groups of organisms. This arrangement of Eukaryota (also called "Eukarya"), Bacteria, and Archaea is called the three-domain system, replacing the traditional two-empire system.
The cells of eukaryotes (left) and prokaryotes (right)
The origin of the eukaryotic cell was a milestone in the evolution of life, since they include all complex cells and almost all multi-cellular organisms. The timing of this series of events is hard to determine; Knoll (2006) suggests they developed approximately 1.6 – 2.1 billion years ago. Some acritarchs are known from at least 1,650 million years ago, and the possible alga Grypania has been found as far back as 2,100 million years ago.
Fossils that are clearly related to modern groups start appearing around 1.2 billion years ago, in the form of a red alga, though recent work suggests the existence of fossilized filamentous algae in the Vindhya basin dating back to 1.6 to 1.7 billion years ago.
Biomarkers suggest that at least stem eukaryotes arose even earlier. The presence of steranes in Australian shales indicates that eukaryotes were present 2.7 billion years ago.
There are many different types of eukaryotic cells, though animals and plants are the most familiar eukaryotes, and thus provide an excellent starting point for understanding eukaryotic structure. Fungi and many protists have some substantial differences, however.
Animal cell
An animal cell is a form of eukaryotic cell that makes up many tissues in animals. The animal cell is distinct from other eukaryotes, most notably plant cells, as they lack cell walls and chloroplasts, and they have smaller vacuoles. Due to the lack of a rigid cell wall, animal cells can adopt a variety of shapes, and a phagocytic cell can even engulf other structures.
There are many different cell types. For instance, there are approximately 210 distinct cell types in the adult human body.
Plant cell
Plant cells are quite different from the cells of the other eukaryotic organisms. Their distinctive features are:
A large central vacuole (enclosed by a membrane, the tonoplast), which maintains the cell's turgor and controls movement of molecules between the cytosol and sap
A primary cell wall containing cellulose, hemicellulose and pectin, deposited by the protoplast on the outside of the cell membrane; this contrasts with the cell walls of fungi, which contain chitin, and the cell envelopes of prokaryotes, in which peptidoglycans are the main structural molecules
The plasmodesmata, linking pores in the cell wall that allow each plant cell to communicate with other adjacent cells; this is different from the functionally analogous system of gap junctions between animal cells.
Plastids, especially chloroplasts that contain chlorophyll, the pigment that gives plants their green color and allows them to perform photosynthesis
Higher plants, including conifers and flowering plants (Angiospermae) lack the flagellae and centrioles that are present in animal cells.
Fungal cell
Fungal cells are most similar to animal cells, with the following exceptions:
A cell wall that contains chitin
Less definition between cells; the hyphae of higher fungi have porous partitions called septa, which allow the passage of cytoplasm, organelles, and, sometimes, nuclei. Primitive fungi have few or no septa, so each organism is essentially a giant multinucleate supercell; these fungi are described as coenocytic.
Only the most primitive fungi, chytrids, have flagella.
Other eukaryotic cells
Eukaryotes are a very diverse group, and their cell structures are equally diverse. Many have cell walls; many do not. Many have chloroplasts, derived from primary, secondary, or even tertiary endosymbiosis; and many do not. Some groups have unique structures, such as the cyanelles of the glaucophytes, the haptonema of the haptophytes, or the ejectisomes of the cryptomonads. Other structures, such as pseudopods, are found in various eukaryote groups in different forms, such as the lobose amoebozoans or the reticulose foraminiferans.
Table 1: Comparison of features of Prokaryotic and Eukaryotic cells
Prokaryotes
Eukaryotes
Typical organisms
Bacteria, archaea
Protists, Fungi, Plants, Animals
Typical size
~ 1–10 µm
~ 10–100 µm (sperm cells, apart from the tail, are smaller)
Type of nucleus
nucleoid region; no real nucleus
real nucleus surrounded by double membrane
DNA
circular (usually)
linear molecules (chromosomes) with histone proteins
RNA-/protein-synthesis
coupled in cytoplasm
RNA-synthesis inside the nucleus protein synthesis in cytoplasm
Ribosomes
50S+30S
60S+40S
Cytoplasmatic structure
very few structures
highly structured by endomembranes and a cytoskeleton
Cell movement
flagella made of flagellin
flagella and cilia containing microtubules; lamellipodia and filopodia containing actin
Mitochondria
none
one to several thousand (though some lack mitochondria)
Chloroplasts
none
in algae and plants
Organization
usually single cells
single cells, colonies, higher multicellular organisms with specialized cells
Structure of a typical animal cellStructure of a typical plant cell
Plant cells are eukaryotic cells that differ in several key respects from the cells of other eukaryotic organisms. Their distinctive features include: A large central vacuole, a water-filled volume enclosed by a membrane known as the tonoplast maintains the cell's turgor, controls movement of molecules between the cytosol and sap, stores useful material and digests waste proteins and organelles.
A cell wall composed of cellulose and hemicellulose, pectin and in many cases lignin, are secreted by the protoplast on the outside of the cell membrane. This contrasts with the cell walls of fungi (which are made of chitin), and of bacteria, which are made of peptidoglycan.
Specialised cell-cell communication pathways known as plasmodesmata, pores in the primary cell wall through which the plasmalemma and endoplasmic reticulum of adjacent cells are continuous.
Plastids, the notables one being the chloroplasts, which contain chlorophyll and the biochemical systems for light harvesting and photosynthesis, but also amyloplasts specialized for starch storage, elaioplasts specialized for fat storage, and chromoplasts specialized for synthesis and storage of pigments. As in mitochondria, which have a genome encoding 37 genes, plastids have their own genomes of about 100-120 unique genes and, it is presumed, arose as prokaryotic endosymbionts living in the cells of an early eukaryotic ancestor of the land plants and algae.
Unlike animal cells, plant cells are stationary.
Cell division by construction of a phragmoplast as a template for building a cell plate late in cytokinesis is characteristic of land plants and a few groups of algae, the notable one being the Charophytes and the Order Trentepohliales.
The sperm of bryophytes have flagellae similar to those in animals, but higher plants, (including Gymnosperms and flowering plants) lack the flagellae and centrioles that are present in animal cells.
Table 2: Comparison of structures between animal and plant cells
Typical animal cell
Typical plant cell
Organelles
Nucleus
Nucleolus (within nucleus)
Rough endoplasmic reticulum (ER)
Smooth ER
Ribosomes
Cytoskeleton
Golgi apparatus
Cytoplasm
Mitochondria
Vesicles
Lysosomes
Centrosome
Centrioles
Nucleus
Nucleolus (within nucleus)
Rough ER
Smooth ER
Ribosomes
Cytoskeleton
Golgi apparatus (dictiosomes)
Cytoplasm
Mitochondria
Plastids and its derivatives
Vacuole(s)
Cell wall
Origin of Eukaryotic organelles and endosymbiotic theoryedit
The endosymbiotic (from the Greek: endo- meaning inside and -symbiosis meaning cohabiting) theory was first articulated by the Russian botanist Konstantin Mereschkowski in 1905. Mereschkowski was familiar with work by botanist Andreas Schimper, who had observed in 1883 that the division of chloroplasts in green plants closely resembled that of free-living cyanobacteria, and who had himself tentatively proposed (in a footnote) that green plants had arisen from a symbiotic union of two organisms. Ivan Wallin extended the idea of an endosymbiotic origin to mitochondria in the 1920s.These theories were initially dismissed or ignored. More detailed electron microscopic comparisons between cyanobacteria and chloroplasts (for example studies by Hans Ris), combined with the discovery that plastids and mitochondria contain their own DNA (which by that stage was recognized to be the hereditary material of organisms) led to a resurrection of the idea in the 1960s.
The endosymbiotic theory was advanced and substantiated with microbiological evidence by Lynn Margulis in a 1967 paper, The Origin of Mitosing Eukaryotic Cells.
In her 1981 work Symbiosis in Cell Evolution she argued that eukaryotic cells originated as communities of interacting entities, including endosymbiotic spirochaetes that developed into eukaryotic flagella and cilia. This last idea has not received much acceptance, because flagella lack DNA and do not show ultrastructural similarities to bacteria or archaea. According to Margulis and Dorion Sagan, "Life did not take over the globe by combat, but by networking" (i.e., by cooperation). The possibility that peroxisomes may have an endosymbiotic origin has also been considered, although they lack DNA. Christian de Duve proposed that they may have been the first endosymbionts, allowing cells to withstand growing amounts of free molecular oxygen in the Earth's atmosphere. However, it now appears that they may be formed de novo, contradicting the idea that they have a symbiotic origin.
It is believed that over millennia these endosymbionts transferred some of their own DNA to the host cell's nucleus during the evolutionary transition from a symbiotic community to an instituted eukaryotic cell (called "serial endosymbiosis"). This hypothesis is thought to be possible because it is known today from scientific observation that transfer of DNA occurs between bacteria species, even if they are not closely related. Bacteria can take up DNA from their surroundings and have a limited ability to incorporate it into their own genome.
Eukaryotes are one of the structurally complex cell type, and by definition are in part organized by smaller interior compartments, that are themselves enclosed by lipid membranes that resemble the outermost cell membrane. The larger organelles, such as the nucleus and vacuoles, are easily visible with the light microscope. They were among the first biological discoveries made after the invention of the microscope.
Not all eukaryotic cells have each of the organelles listed below. Exceptional organisms have cells which do not include some organelles that might otherwise be considered universal to eukaryotes (such as mitochondria).[22] There are also occasional exceptions to the number of membranes surrounding organelles, listed in the tables below (e.g., some that are listed as double-membrane are sometimes found with single or triple membranes). In addition, the number of individual organelles of each type found in a given cell varies depending upon the function of that cell.
Major eukaryotic organelles
Organelle
Main function
Structure
Organisms
Notes
Chloroplast (plastid)The simplified internal structure of a chloroplast
photosynthesis
double-membrane compartment
plants, protists (rare kleptoplastic organisms)
has some genes; theorized to be engulfed by the ancestral eukaryotic cell (endosymbiosis)
translation and folding of new proteins (rough endoplasmic reticulum), expression of lipids (smooth endoplasmic reticulum)
single-membrane compartment
all eukaryotes
rough endoplasmic reticulum is covered with ribosomes, has folds that are flat sacs; smooth endoplasmic reticulum has folds that are tubular
Golgi apparatusDiagram of secretory process from endoplasmic reticulum (orange) to Golgi apparatus (pink). 1. Nuclear membrane; 2. Nuclear pore; 3. Rough endoplasmic reticulum (RER); 4. Smooth endoplasmic reticulum (SER); 5. Ribosome attached to RER; 6. Macromolecules; 7. Transport vesicles; 8. Golgi apparatus; 9. Cis face of Golgi apparatus; 10. Trans face of Golgi apparatus; 11. Cisternae of lipids
sorting and modification of proteins
single-membrane compartment
all eukaryotes
cis-face (convex) nearest to rough endoplasmic reticulum; trans-face (concave) farthest from rough endoplasmic reticulum
energy production (house), Mitochondria are self-replicating organelles that occur in various numbers, shapes, and sizes in the cytoplasm of all eukaryotic cells.
double-membrane compartment
most eukaryotes
has some DNA; theorized to be engulfed by an ancestral eukaryotic cell (endosymbiosis)
VacuoleThe central vacuole within the cytoplasm of a plant cell
NucleusHeLa cells stained for DNA with the BlueHoechst dye. The central and rightmost cell are in interphase, thus their entire nuclei are labeled. On the left, a cell is going through mitosis and its DNA has condensed ready for division.
It houses the cell's chromosomes, and is the place where almost all DNA replication, RNA transcription take place
Mitochondria and chloroplasts, which have double-membranes and their own DNA, are believed to have originated from incompletely consumed or invading prokaryotic organisms, which were adopted as a part of the invaded cell. This idea is supported in the Endosymbiotic theory.
Minor eukaryotic organelles and cell components
Organelle/Macromolecule
Main function
Structure
Organisms
AcrosomeA picture of a spermazoa cell with its acrosome colored in red
helps spermatoza fuse with ovum
single-membrane compartment
many animals
AutophagosomeAutophagosomes labeled by a fluorescent marker.
vesicle which sequesters cytoplasmic material and organelles for degradation
(A) Electron micrograph of Halothiobacillus neapolitanus cells, arrows highlight carboxysomes. (B) Image of intact carboxysomes isolated from H. neapolitanus. Scale bars are 100 nm.[24]
Prokaryotes are not as structurally complex as eukaryotes, and were once thought not to have any internal structures enclosed by lipid membranes. In the past, they were often viewed as having little internal organization; but, slowly, details are emerging about prokaryotic internal structures. An early false turn was the idea developed in the 1970s that bacteria might contain membrane folds termed mesosomes, but these were later shown to be artifacts produced by the chemicals used to prepare the cells for electron microscopy.[25]
However, more recent research has revealed that at least some prokaryotes have microcompartments such as carboxysomes. These subcellular compartments are 100 - 200 nm in diameter and are enclosed by a shell of proteins.[26] Even more striking is the description of membrane-bound magnetosomes in bacteria,[27][28] as well as the nucleus-like structures of the Planctomycetes that are surrounded by lipid membranes.[29]
Macromolecules which are present in the cell membraneedit
Cell membranes contain a variety of biological molecules, notably lipids and proteins. Material is incorporated into the membrane, or deleted from it, by a variety of mechanisms:
Fusion of intracellular vesicles with the membrane (endocytosis) not only excretes the contents of the vesicle but also incorporates the vesicle membrane's components into the cell membrane. The membrane may form blebs around extracellular material that pinch off to become vesicles (exocytosis).
If a membrane is continuous with a tubular structure made of membrane material, then material from the tube can be drawn into the membrane continuously.
Although the concentration of membrane components in the aqueous phase is low (stable membrane components have low solubility in water), there is an exchange of molecules between the lipid and aqueous phases.
Lipids
The cell membrane consists of three classes of amphipathic lipids: phospholipids, glycolipids, and cholesterols. The amount of each depends upon the type of cell, but in the majority of cases phospholipids are the most abundant. In RBC studies, 30% of the plasma membrane is lipid.
The fatty chains in phospholipids and glycolipids usually contain an even number of carbon atoms, typically between 16 and 20. The 16- and 18-carbon fatty acids are the most common. Fatty acids may be saturated or unsaturated, with the configuration of the double bonds nearly always cis. The length and the degree of unsaturation of fatty acid chains have a profound effect on membrane fluidity as unsaturated lipids create a kink, preventing the fatty acids from packing together as tightly, thus decreasing the melting temperature (increasing the fluidity) of the membrane. The ability of some organisms to regulate the fluidity of their cell membranes by altering lipid composition is called homeoviscous adaptation.
The entire membrane is held together via non-covalent interaction of hydrophobic tails, however the structure is quite fluid and not fixed rigidly in place. Under physiological conditions phospholipid molecules in the cell membrane are in the liquid crystalline state. It means the lipid molecules are free to diffuse and exhibit rapid lateral diffusion along the layer in which they are present. However, the exchange of phospholipid molecules between intracellular and extracellular leaflets of the bilayer is a very slow process. Lipid rafts and caveolae are examples of cholesterol-enriched microdomains in the cell membrane.
In animal cells cholesterol is normally found dispersed in varying degrees throughout cell membranes, in the irregular spaces between the hydrophobic tails of the membrane lipids, where it confers a stiffening and strengthening effect on the membrane.
Lipid vesicles or liposomes are circular pockets that are enclosed by a lipid bilayer. These structures are used in laboratories to study the effects of chemicals in cells by delivering these chemicals directly to the cell, as well as getting more insight into cell membrane permeability. Lipid vesicles and liposomes are formed by first suspending a lipid in an aqueous solution then agitating the mixture through sonication, resulting in a uniformly circular vesicle. By measuring the rate of efflux from that of the insideof the vesicle to the ambient solution, allows researcher to better understand membrane permeability. Vesicles can be formed with molecules and ions inside the vesicle by forming the vesicle with the desired molecule or ion present in the solution. Proteins can also be embedded into the membrane through solubilizing the desired proteins in the presence of detergents and attaching them to the phospholipids in which the liposome is formed. These provide researchers with a tool to examine various membrane protein functions.
Carbohydrates
Plasma membranes also contain carbohydrates, predominantly glycoproteins, but with some glycolipids (cerebrosides and gangliosides). For the most part, no glycosylation occurs on membranes within the cell; rather generally glycosylation occurs on the extracellular surface of the plasma membrane.
The glycocalyx is an important feature in all cells, especially epithelia with microvilli. Recent data suggest the glycocalyx participates in cell adhesion, lymphocyte homing, and many others.
The penultimate sugar is galactose and the terminal sugar is sialic acid, as the sugar backbone is modified in the golgi apparatus. Sialic acid carries a negative charge, providing an external barrier to charged particles.
Proteins
Proteins within the membrane are key to the functioning of the overall membrane. These proteins mainly transport chemicals and information across the membrane. Every membrane has a varying degree of protein content. Proteins can be in the form of peripheral or integral.
The cell membrane plays host to a large amount of protein that is responsible for its various activities. The amount of protein differs between species and according to function, however the typical amount in a cell membrane is 50%.These proteins are undoubtedly important to a cell: Approximately a third of the genes in yeast code specifically for them, and this number is even higher in multicellular organisms.
The cell membrane, being exposed to the outside environment, is an important site of cell-cell communication. As such, a large variety of protein receptors and identification proteins, such as antigens, are present on the surface of the membrane. Functions of membrane proteins can also include cell-cell contact, surface recognition, cytoskeleton contact, signaling, enzymatic activity, or transporting substances across the membrane.
Most membrane proteins must be inserted in some way into the membrane. For this to occur, an N-terminus "signal sequence" of amino acids directs proteins to the endoplasmic reticulum, which inserts the proteins into a lipid bilayer. Once inserted, the proteins are then transported to their final destination in vesicles, where the vesicle fuses with the target membrane.
Some theorists suggest that the atmosphere of the early Earth may have been chemically reducing in nature, composed primarily of methane (CH4), ammonia (NH3), water (H2O), hydrogen sulfide (H2S), carbon dioxide (CO2) or carbon monoxide (CO), and phosphate (PO43-), with molecular oxygen (O2) and ozone (O3) either rare or absent.
The sequence of chemical events that led to the first nucleic acids is not known.
In such a reducing atmosphere, electrical activity can catalyze the creation of certain basic small molecules (monomers) of life, such as amino acids. This was demonstrated in the Miller–Urey experiment by Stanley L. Miller and Harold C. Urey in 1953.
Phospholipids (of an appropriate length) can spontaneously form lipid bilayers, a basic component of the cell membrane.
The polymerization of nucleotides into random RNA molecules might have resulted in self-replicating ribozymes.
Synthesized proteins might then outcompete ribozymes in catalytic ability, and therefore become the dominant biopolymer, relegating nucleic acids to their modern use, predominantly as a carrier of genomic information.
1632–1723: Antonie van Leeuwenhoek teaches himself to grind lenses, builds a microscope and draws protozoa, such as Vorticella from rain water, and bacteria from his own mouth.
1665: Robert Hooke discovers cells in cork, then in living plant tissue using an early microscope.
1839: Theodor Schwann and Matthias Jakob Schleiden elucidate the principle that plants and animals are made of cells, concluding that cells are a common unit of structure and development, and thus founding the cell theory.
The belief that life forms can occur spontaneously (generatio spontanea) is contradicted by Louis Pasteur (1822–1895) (although Francesco Redi had performed an experiment in 1668 that suggested the same conclusion).
1855: Rudolf Virchow states that cells always emerge from cell divisions (omnis cellula ex cellula).
1931: Ernst Ruska builds first transmission electron microscope (TEM) at the University of Berlin. By 1935, he has built an EM with twice the resolution of a light microscope, revealing previously unresolvable organelles.
1953: Watson and Crick made their first announcement on the double-helix structure for DNA on February 28.
1981: Lynn Margulis published Symbiosis in Cell Evolution detailing the endosymbiotic theory.
Evidence which support endosymbiotic theory
Mitochondria and plastids are formed only through a process similar to binary fission. In some algae, such as Euglena, the plastids can be destroyed by certain chemicals or prolonged absence of light without otherwise affecting the cell. In such a case, the plastids will not regenerate.
They are surrounded by two or more membranes, and the innermost of these shows differences in composition from the other membranes of the cell. They are composed of a peptidoglycan cell wall characteristic of a bacterial cell.
Both mitochondria and plastids contain DNA that is different from that of the cell nucleus and that is similar to that of bacteria (in being circular in shape and in its size).
DNA sequence analysis and phylogenetic estimates suggest that nuclear DNA contains genes that probably came from plastids.
These organelles' ribosomes are like those found in bacteria (70S).
Proteins of organelle origin, like those of bacteria, use N-formylmethionine as the initiating amino acid.
Much of the internal structure and biochemistry of plastids, for instance the presence of thylakoids and particular chlorophylls, is very similar to that of cyanobacteria.
Phylogenetic estimates constructed with bacteria, plastids, and eukaryotic genomes also suggest that plastids are most closely related to cyanobacteria.
Mitochondria have several enzymes and transport systems similar to those of bacteria.
Some proteins encoded in the nucleus are transported to the organelle, and both mitochondria and plastids have small genomes compared to bacteria. This is consistent with an increased dependence on the eukaryotic host after forming an endosymbiosis. Most genes on the organellar genomes have been lost or moved to the nucleus. Most genes needed for mitochondrial and plastid function are located in the nucleus. Many originate from the bacterial endosymbiont.
Plastids are present in very different groups of protists, some of which are closely related to forms lacking plastids. This suggests that if chloroplasts originated de novo, they did so multiple times, in which case their close similarity to each other is difficult to explain.
Many of these protists contain "primary" plastids that have not yet been acquired from other plastid-containing eukaryotes.
Among eukaryotes that acquired their plastids directly from bacteria (known as Primoplantae), the glaucophyte algae have chloroplasts that strongly resemble cyanobacteria. In particular, they have a peptidoglycan cell wall between the two membranes.
Mitochondria and plastids are similar in size to bacteria.
↑Thompson WR, Murray BG, Khare BN, Sagan C (1987). "Coloration and darkening of methane clathrate and other ices by charged particle irradiation: applications to the outer solar system". Journal of geophysical research. 92 (A13): 14933–47. doi:10.1029/JA092iA13p14933. PMID11542127. {{cite journal}}: Unknown parameter |month= ignored (help)CS1 maint: multiple names: authors list (link)
↑Hill HG, Nuth JA (2003). "The catalytic potential of cosmic dust: implications for prebiotic chemistry in the solar nebula and other protoplanetary systems". Astrobiology. 3 (2): 291–304. doi:10.1089/153110703769016389. PMID14577878.
↑Balm SP, Hare J.P., Kroto HW (1991). "The analysis of comet mass spectrometric data". Space Science Reviews. 56: 185–9. doi:10.1007/BF00178408.{{cite journal}}: CS1 maint: multiple names: authors list (link)
↑Miller, Stanley L. (1959). "Organic Compound Synthesis on the Primitive Earth". Science. 130 (3370): 245. doi:10.1126/science.130.3370.245. PMID13668555. {{cite journal}}: Unknown parameter |coauthors= ignored (|author= suggested) (help); Unknown parameter |month= ignored (help) Miller states that he made "A more complete analysis of the products" in the 1953 experiment, listing additional results.
↑A. Lazcano, J. L. Bada (2004). "The 1953 Stanley L. Miller Experiment: Fifty Years of Prebiotic Organic Chemistry". Origins of Life and Evolution of Biospheres. 33 (3): 235–242. doi:10.1023/A:1024807125069. PMID14515862. {{cite journal}}: Unknown parameter |month= ignored (help)
↑Oró J, Kimball AP (1961). "Synthesis of purines under possible primitive earth conditions. I. Adenine from hydrogen cyanide". Archives of biochemistry and biophysics. 94: 217–27. doi:10.1016/0003-9861(61)90033-9. PMID13731263. {{cite journal}}: Unknown parameter |month= ignored (help)
↑Oró J, Kamat SS (1961). "Amino-acid synthesis from hydrogen cyanide under possible primitive earth conditions". Nature. 190: 442–3. doi:10.1038/190442a0. PMID13731262. {{cite journal}}: Unknown parameter |month= ignored (help)
↑Oró J (1967). Fox SW (ed.). Origins of Prebiological Systems and of Their Molecular Matrices. New York Academic Press. p. 137.
↑Ryter A (1988). "Contribution of new cryomethods to a better knowledge of bacterial anatomy". Ann. Inst. Pasteur Microbiol. 139 (1): 33–44. doi:10.1016/0769-2609(88)90095-6. PMID3289587.
↑Kerfeld CA, Sawaya MR, Tanaka S, Nguyen CV, Phillips M, Beeby M, Yeates TO (2005). "Protein structures forming the shell of primitive bacterial organelles". Science. 309 (5736): 936–8. doi:10.1126/science.1113397. PMID16081736. {{cite journal}}: Unknown parameter |month= ignored (help)CS1 maint: multiple names: authors list (link)
↑Komeili A, Li Z, Newman DK, Jensen GJ (2006). "Magnetosomes are cell membrane invaginations organized by the actin-like protein MamK". Science. 311 (5758): 242–5. doi:10.1126/science.1123231. PMID16373532.{{cite journal}}: CS1 maint: multiple names: authors list (link)
↑Scheffel A, Gruska M, Faivre D, Linaroudis A, Plitzko JM, Schüler D (2006). "An acidic protein aligns magnetosomes along a filamentous structure in magnetotactic bacteria". Nature. 440 (7080): 110–4. doi:10.1038/nature04382. PMID16299495.{{cite journal}}: CS1 maint: multiple names: authors list (link)










.svg)
















